This manuscript was originally presented at the NACF, MARDI, IFOAM-Asia, and FFTC workshop titled “Technology, Implementation, and Policy Review of Biofertilizers and Biopesticides in the Asian and Pacific Region,” held at the NACF Headquarters in Seoul, Korea, on May 8–10, 2024.
ABSTRACT
With the increase in world population, the demography of humans is estimated to be exceeded, and it has become a major challenge to provide an adequate amount of healthy food globally. The use of chemical fertilizers and fungicides causes the plant to grow efficiently and rapidly to meet the food demand. But the drawbacks of using a higher quantity of chemicals are environmental pollution, changes in the soil ecology, physiochemical composition, decreasing agricultural yields leading to several health hazards. Climatic factors such as abiotic and biotic stresses like soil salinity, drought, wind, raising temperatures, heavy metals, waterlogging, and different weeds and phytopathogens like bacteria, viruses, fungi, and nematodes which attack plants, reducing crop productivity and quality. In recent years, the growing adoption of organic farming has resulted in a more sustainable approach towards agriculture as it emphasises on adopting environmentally friendly practices to ensure food security while protecting and nurturing soil health. Various strategies to make organic and sustainable agriculture, one of the key factors being the use of biofertilizers (PGPR) to provide optimum nutrients to crop and enhance soil health. There is a shift toward the use of biofertilizers due to all these facts, which provide nutrition through natural processes like zinc, potassium and phosphorus solubilization, nitrogen fixation, production of hormones, siderophore, various hydrolytic enzymes and protect the plant from different plant pathogens and stress conditions. They are made up of physiologically active microbes, such as bacteria, algae, and fungal species, that help to improve, preserve, and transform nutrients from an unusable to a useful form. There are various ways to introduce biofertilizers, such as root dipping, in-furrow soil application and inoculating onto seeds with a liquid or dry formulation. The recently emerging techniques such as nano-biofertilizer and biofilm biofertilizer enable crops to become more resistant toward environmental inhibitors and pathogenic components. The advanced microbial inoculants by formulating various plant growth-promoting rhizobacteria (PGPR) to combat plant diseases and environmental stress is getting more popular in recent days. They are often regarded as a renewable source of plant nutrients and promote soil health and have become a vital constituent in an integrated crop and soil management systems. The demand for biofertilizers in Asia-Pacific is anticipated to increase throughout the forecast period, with a CAGR of 8.6%. This review critically examines the current state-of-art on use of PGPR’s in different crop in maintaining soil fertility and enhancing crop productivity. It is believed that acquisition of advanced knowledge of plant-PGPR interactions, bioengineering of microbial communities to improve the performance of PGPR under field conditions, will help in devising strategies for sustainable, environment-friendly and climate smart agricultural technologies to deliver short and long terms solutions for improving crop productivity to feed the world in a more sustainable manner.
Keywords: Biofertilizers, Sustainable crop production, Bioactive hormones, Plant growth promotion, Microbial inoculants, PGPR
INTRODUCTION
In a world where global food security is increasingly threatened, adopting sustainable and environmentally friendly Bio-Agri Inputs approaches to safeguard our agricultural crops is of paramount importance. Addressing these pressing issues necessitates a paradigm shift in agricultural practices, promotion of environmentally friendly alternatives for the reduction of chemical inputs is a mandatory approach. In this context, leveraging the interactions between plants and beneficial microorganisms, such as plant growth-promoting rhizobacteria (PGPR), has emerged as a promising strategy and in this context, we refer them as biofertilizers. PGPR, a diverse group of soil-dwelling bacteria, are renowned for their multifaceted roles in enhancing plant growth, improving nutrient uptake, and conferring resistance to various biotic and abiotic stresses. They have the potential to serve as biofertilizers and or biocontrol agents which are substitutes for undesirable chemicals and pesticides, promoting sustainable healthy practices for sustainable and organic agriculture. Over the years, the research has uncovered an intriguing facet of the plant-PGPR interaction, namely, the phenomenon of plant defence priming. Biofertilizers are biologically active formulations comprising beneficial live microbes (PGPR) in economical carrier materials, which when applied to the soil, plant, and seed improve plant growth and development by enhancing the supply of plant growth-promoting substances or nutrients. The term “biofertilizer” has been defined in different ways during the past 20 years, which derives from the improved understanding of the relationships occurring between the rhizosphere microorganisms and the plant. Finally, there is a common consensus among many researchers include ours is that “a substance which contains living microorganisms which, when applied to seed, plant surfaces, or soil, colonizes the rhizosphere or the interior of the plant and promotes growth by increasing the supply or availability of primary nutrients to the host plant”.
Plant biostimulants are becoming popular in the agricultural market as products to increase growth, health, and/or yield of crop plants. The challenge is that there are many types of biostimulants, each with unique mechanisms of action, and these dynamics make it difficult to optimize product placement into grower systems. A global assessment of the structure and function of the crop microbiome is urgently needed for the development of effective and rationally designed microbiome technologies for sustainable agriculture. Such an effort will provide new knowledge on the key ecological and evolutionary interactions between plant species and their microbiomes that can be harnessed for increasing agriculture productivity. Hower, in this review, whatever the term may be, we will be focused on PGPR as a biofertilizer which includes as bioinoculants, biostimulants, biofilm and plant microbiome for ease of understanding.
The groups of microbes mostly used for inoculant production are the arbuscular mycorrhizal (AM) fungi, PGPR and nitrogen-fixing rhizobia (Malusà, et al., 2012). Most soils contain extremely elevated levels of bacteria (often estimated to be ~108–109 bacterial cells per gram of soil). Some of these bacteria are PGPB, some are phytopathogens and some do not have any discernible effect on plant growth. The knowledge of the physiological traits and mechanisms that are used by PGPR to promote plant growth has developed over the past 20–30 years. In fact, while only a small number of PGPR have been commercialized as adjuncts to agricultural practice, both the number of commercialized strains and the amount of farmland devoted to using this approach continue to increase with each succeeding year.
All land plants host a microbiome composed of bacteria, fungi, oomycetes, viruses, archaea, and protists. These organisms primarily colonize the root environment, especially the rhizosphere (the soil portion immediately adjacent to the root), the rhizoplane (intended as the root surface), and to a lesser extent the leaf surface (phyllosphere), seed (spermosphere), and internal (endosphere) plant environments. Therefore, microbiomes associated with plants show a different degree of intimacy with the plant: while epiphytic microorganisms can live externally, endophytic microorganisms efficiently colonize the internal plant tissues. In this context, plants may be viewed as superorganisms that include their microbiomes, which provide several unique functions and features. However, notwithstanding an abundance of recent studies, the knowledge on plant-associated microbiomes and their detailed effects on crop productivity, growth, health, and disease are still scanty. Plants can take advantage of the rhizosphere microbiome by selectively inducing activities within microorganisms that support their development and survival. In this way, rhizodeposition shapes the plant-associated microbiome, which in turn, can affect both plant metabolic pathways and exudate release. Based on this knowledge, the existence of crosstalk among the host–microbiome and metabolome, where plants perceive and understand the chemical communication among microbes and vice-versa. By considering the plants as an holobiont communicating with the external environment, that in turn affects the interrelationships occurring in the holobiont itself, where molecules affecting rhizosphere interactions and plant health can be degraded, used, and modified by the members of the rhizosphere microbiome.
Until now, most of the knowledge and understanding of how PGPR promote plant growth has included studies of isolated bacteria examined in controlled laboratory conditions that have not considered the range of organisms and abiotic conditions that affect how these bacteria function in soil. In the soil, PGPR typically acts together in groups or consortia, with the consortia being responsible for facilitating plant growth, that is, different bacteria within the consortia satisfy different plant needs. The attraction of both individual PGPR, as well as bacterial consortia of PGPR, to a particular plant is dictated, in part, by the range and concentration of small molecule root exudates produced by a particular plant. Interestingly, even though the microbiota in most microbiomes contains both PGPR and deleterious microorganisms plants attract beneficial PGPR and not phytopathogens. Thus, despite the presence of phytopathogens in many microbiomes, the beneficial PGPR often protects plants from the deleterious effects of phytopathogens.
The world's population is continually growing, and projections indicate that by 2050, there will be approximately 9.7 billion people on the planet. This quick expansion is closely linked to extensive urbanization, industrialization, and agricultural output (Macik et al., 2020). Numerous hybrid crop varieties have been developed and are still being developed to meet the demands of the expanding population.
The efficient application of chemical fertilizers to crops yielding undeniable benefits for crop growth, yield, and quality as well as financial gain for farmers. Unfortunately, because of the greater quantities of synthetic fertilizers are being used, leads to soil, water, and air pollution which could be a serious threat to the ecosystem. These synthetic chemicals are being applied carelessly causes their accumulation below ground. This leads to adverse changes in the structure, fertility, and water-holding capacity of the soil. In-addition, overuse of synthetic fertilizers is also strongly associated with the greenhouse effect and eutrophication of water bodies. Extended usage of mineral fertilizers can cause the soil's nutrient level to decrease and increase crop susceptibility to several diseases (Macik et al., 2020).
For better yields, farmers applying lot of chemical fertilizers that can increase yield but damage the soil strength and biota. To overcoming these problems recent agricultural practices mainly utilization of biofertilizers such as Rhizobium strains, PSB (phosphate solubilizing bacteria), VAM (Vascular Arbuscular Mycorrhiza), Cyanobacteria and various other microorganism plays a vital role on crops in increasing nutrients uptake by fixation of nutrients by forming nodules in roots that helps in mineralization process that can convert the unavailable form of nutrients to available form that can easily uptake by plant. These biofertilizers are mostly host specific that can symbiont with pulse crop and boost the growth and yield attributes (Santhosh and Sandeep, 2020).
TheGeenera Gigaspora, Glomus, Bacillus, Pseudomonas, Azotobacter, Azospirillum, Bradyrhizobium and Rhizobium, are commonly used as PGPR (Adeleke, 2019, Thomas and Singh, 2019). These PGPR play essential roles in soil ecosystem functions such as restoring and preserving soil nutrient richness. Several studies have been conducted on PGPR and their various benefits in sustainable agriculture, especially in promoting plant nutrition (García-Fraile, 2015, Lawal and Babalola, 2014). A more environmentally friendly option to chemical fertilizers is PGPR or microbial inoculants. They boost agricultural yields while conserving the lithosphere and benefiting the biosphere by preventing soil, water, and air pollution and eutrophication. The use of PGPR containing strains of beneficial microorganisms is one strategy used in sustainable farming to reduce issues brought on by overuse of mineral fertilizers (Mishra and Dash, 2014).
TYPES OF PGPR
PGPR can improve nutrient absorption and promote overall plant growth. These PGPR form symbiotic relationships with plants or fix atmospheric nitrogen, making it accessible to plants in a usable form. There are diverse types of PGPR, each serving a specific purpose on crops:
Nitrogen-fixing PGPR: These contain nitrogen-fixing bacteria, such as Rhizobium, Azotobacter, and Azospirillum, which convert atmospheric nitrogen into a form that plants can utilize.
Phosphorus-solubilizing PGPR: These consist of bacteria and fungi, such as Bacillus and Pseudomonas, which help solubilize insoluble phosphates in the soil, making phosphorus more available to plants.
Potassium-mobilizing PGPR: These contain microorganisms that enhance the availability of potassium in the soil, improving the plant's ability to absorb this essential nutrient.
Symbiotic PGPR: These involve the use of mycorrhizal fungi or nitrogen-fixing bacteria that form symbiotic relationships with plant roots, aiding in nutrient absorption and overall plant health.
For ease of understanding, we categorized them based on their mode of action in the following sub-headings which are as follows:
Nutrient enhancement: PGPR that can fix atmospheric nitrogen and solubilize phosphorus, making these essential nutrients available to plants which could enhance soil fertility, healthier plant growth and their crop yields.
Reduced dependency on fertilizers: The application of PGPR reduces the reliance on synthetic or chemical fertilizers, which can have detrimental effects on soil health and the environment over time. Biofertilizers can offer a more sustainable and eco-friendlier alternative by promoting natural nutrient cycles.
Environmental sustainability: PGPR contributes to the conservation of soil, water, and air quality. These helps maintain the balance of ecosystems by minimizing the negative impacts associated with the excessive use of chemical fertilizers, such as soil degradation, water pollution, and greenhouse gas emissions.
Promotion of microbial activity: PGPR introduces beneficial microorganisms into the soil, fostering a healthy microbial community around plant root ecosystem. These beneficial microbes play a crucial role in nutrient cycling, disease control attributes, and overall healthy soil ecosystem functioning.
Biodiversity conservation: By promoting sustainable farming practices, PGPR contributes to the conservation of biodiversity. Reduced dependence on chemical inputs helps maintain a more balanced and diverse ecosystem in and around agricultural areas.
Improved soil health: The activities of PGPR can enhance soil structure by promoting the formation of aggregates. This improves water retention, aeration, and drainage in the soil, leading to better overall soil quality.
Cost-effectiveness: In modern agriculture, PGPR can be cost-effective for farmers for sustainable agriculture. Though the initial investment might be higher, the reduced need for chemical fertilizers, coupled with their improved soil fertility and crop yields, can lead to overall economic returns to farmers.
Plant tolerance: Some PGPR that can enhance a plant's resistance to various environmental stresses, such as drought, salinity, and certain diseases. This can contribute to increased crop resilience and adaptability to changing climatic conditions.
PGPR MARKET TRENDS
Some of the biofertilizers market trends are:
Growing demand for organic agriculture: The increasing global demand for organic produce has been a significant driver for the PGPR market. Consumers are more conscious of the environmental impact of conventional farming practices, leading to a shift towards organic and sustainable agriculture.
Government initiatives and regulations: Most of the governments around the globe have been promoting the use of PGPR through subsidies, incentives, and regulations. This support is aimed at encouraging farmers to adopt eco-friendly and sustainable agricultural practices.
Advancements in PGPR strains: Current research and development in microbiology have led to the identification and isolation of more effective microbial strains for PGPR production. Improved strains contribute to better nutrient availability and overall plant health.
Education in PGPR: Farmers and agricultural practitioners are becoming more aware of the benefits of PGPR. Educational programs and initiatives by agricultural extension services contribute to the dissemination of knowledge regarding the advantages of using biobased PGPR.
Compatibility with conventional chemical fertilizers: PGPR are increasingly being integrated into conventional fertilizer practices. This integrated approach helps optimize nutrient management, reduce dependency on chemical fertilizers, and enhance soil health.
Bio-technological innovations in PGPR production: Bio-technological innovations in the production of PGPR have led to increased efficiency and cost-effectiveness. Improved manufacturing processes and packaging methods contribute to the wider adoption of PGPR.
Focus on Nitrogen-fixing PGPR: Nitrogen-fixing bacteria, such as Rhizobium and Azotobacter, are gaining prominence due to their role in enhancing nitrogen availability in the soil. This is particularly crucial, as nitrogen is a vital nutrient for plant growth.
Market opportunities and partnerships: The PGPR industry has witnessed various mergers, acquisitions, and strategic partnerships. Such collaborations help companies broaden their product portfolios, enhance research capabilities, and expand their market reach.
PGPR in climate resilience: The environmental benefits of PGPR, such as reduced greenhouse gas emissions and improved soil structure, align with global sustainability goals. As climate change concerns grow, the emphasis on sustainable agricultural practices is likely to drive the adoption of PGPR.
APPLICATION OF PGPR FOR SUSTAINABLE AGRICULTURE
Seed application: PGPR can be applied directly to seeds before planting. This mode ensures that the beneficial microorganisms establish contact with the emerging roots in the preliminary stages of plant growth.
Soil Application: PGPR can be mixed with soil before sowing seeds or transplanting seedlings. This allows the PGPR to colonize the root zone, promoting a symbiotic relationship with the plants.
Seedling drench or dip: Seedlings can be immersed in a PGPR solution before transplanting. This method ensures that the young plants are inoculated with beneficial microorganisms, fostering a favorable environment for root development. A liquid PGPR solution can be poured or sprayed directly onto the roots of established plants. This method facilitates the direct application of beneficial PGPR to the root system, enhancing nutrient uptake.
Foliar spray: PGPR can be applied as a foliar spray onto the leaves of plants. This spay allows for the absorption of nutrients through the stomata on the leaf surface, providing a supplemental nutrient source to support plant growth.
Drip irrigation: PGPR can be injected into irrigation systems for controlled and uniform distribution. This system ensures that plants receive a consistent supply of beneficial microorganisms along with water.
Integration with low dose fertigation: This method involves the integration of PGPR into conventional low dose fertilizer application practices. It combines fertilization with irrigation, allowing for the simultaneous delivery of nutrients and beneficial microorganisms to the root zone.
Application with compost: PGPR can be incorporated into composting processes. Adding beneficial microorganisms during composting enhances the microbial diversity and nutrient content of the final compost, which can then be applied to the soil.
Application in hydroponics: In soilless growing systems like aquaponics and hydroponics, PGPR can be introduced to the nutrient solution. This application provides a source of beneficial microorganisms to support plant growth in these controlled environments.
The formulation of PGPR can consist of one or more distinct microbial strains, such as AM, Bacillus, Pseudomonas, Rhizobium, Acetobacter, Azotobacter, Rhizobium and other PGPR strains. The PGPR can be classified into different groups.
NITROGEN FIXING PGPR
An element necessary for life, nitrogen (N) is a part of amino acids, nucleic acids, chlorophyll, and the energy-transfer molecule adenosine triphosphate (ATP). Organic N is one of the main sources of nitrogen in soils. It must undergo microbial mineralization i.e., through processes like nitrification and ammonification, to be transformed into inorganic N that is available to plants. The main reservoir of nitrogen, however, is found in the atmosphere as N2, which is only made available via biological nitrogen-fixation and is not utilized by plants directly. The energy-intensive process of biological nitrogen fixation involves the conversion of atmospheric N2 to ammonia (NH3) by the enzyme nitrogenase. This is a readily assimilable form for both plants and microorganisms. Nitrogenases are present in a relatively few diverse classes of microorganisms known as diazotrophs (N2-fixing), which includes free-living bacteria, symbiotic bacteria, and archaea. Multiple bacterial strains, such as aerobic (Azotobacter), anaerobic (Clostridium), facultatively anaerobic (Klebsiella), or heterotrophs; anoxygenic (Rhodobacter) or oxygenic (Anabaena) phototrophs and chemolithotrophs (Leptospirillum ferrooxidans), can carry out nitrogen fixation with varying physiological requirements. Diazotrophs inhabit a variety of soil and aquatic habitats. They can establish symbiotic relationships with cyanobacteria, and actinorhizal associations with woody plants, symbiotic relationships with legumes that result in the production of root nodules. BNF permits the replenishment of the total nitrogen contentand fixed nitrogen controls crop productivity and growth.
The Rhizobiaceae family of bacteria, which includes Rhizobium, Bradyrhizobium, Sinorhizobium, Azorhizobium, Mesorhizobium, and Sinorhizobium, are the most studied group of symbiotic N2-fixing organisms. By forming nodules on the roots or stems of legumes, they can form symbiotic associations. The benefit of these nodules for N2-fixation is that bacteroids' nitrogenases are protected from atmosphere O2. Given that oxygen is a negative regulator of nif gene expression and inhibits nitrogenase activity, the quantity of oxygen plays a significant role in controlling the amount of N that is fixed. A considerable increase in plant biomass and grain yield was seen in a variety of legume crops after efficient rhizobia strains were inoculated. Rhizobium strain inoculation of soybeans resulted in an overall increase in growth and yield components, including the number of branches yielding pods per plant, the total number of pods per plant, and the number of seeds per pod (Mfilinge et al., 2014). Bourion et al., (2018) reported that when R. leguminosarum strains inoculated into pea and lentil seeds, nodulation, shoot/root weight, and pea seed production were all increased. Legume plants are less reliant on chemical fertilizers than non-leguminous plants because of the nitrogen fixation process that Rhizobium performs.
Several heterotrophic free-living diazotrophic microorganisms, including cyanobacteria, Azotobacter sp. and Azospirillum sp., can fix atmospheric N2 in the rhizosphere and bulk soil. In non-legume crops, free-living diazotrophs play a crucial role in N acquisition. enhanced yields of many crops, including tomato, eggplant, pepper, cotton, sunflower, carrot, oak, and sugar beet, have been observed, for instance, along with enhanced yields of rice, corn, and wheat (Garcha and Maan, 2017). According to Jain et al., (2021) findings, seven of the 24 Azotobacter strains that were identified demonstrated phosphate solubilization, while allthe 24 strains produced ammonia, siderophores, HCN, and IAA. Certain strains of cyanobacteria have been shown to boost the growth and yield of radish, maize, tomato, cotton, barley, and oats. Several legumes showed significantly enhanced nodule number, nitrogen fixation, plant biomass, and total nitrogen contents when compared to uninoculated plants or a single Rhizobium inoculation. The most well-known free-living diazotrophs, Azospirillum species have been proved to improve N availability and acquisition in addition to performing a number other PGP activity. Azospirillum species associates with plant roots and synthesize various compounds involved in plant growth promotion, for instance IAA, gibberellins and cytokinin (Pereg et al., 2016) are shown in Table 1.
Table 1. Types of nitrogen fixing PGPR strains.
|
Types of biofertilizer
|
PGPR strain used
|
Plant growth promoting activity
|
Effect on plant productivity parameters
|
|
Nitrogen-fixing
bacteria
|
Azotobacter sp. strain Avi2 (MCC 3432)
|
Nitrogen fixation, production of
IAA, siderophore
|
Improved vegetative and reproductive growth in
rice
|
|
Azotobacter chroococcum, A. vinelandii
|
Nitrogen fixation, P
solubilization, production of
NH3, HCN, IAA
|
Increased shoot and root length, leaf and rootnumber, chlorophyll content of maize
|
|
Herbaspirillum seropedicae (strain
ZAE94)
|
Nitrogen fixation, production of
siderophore, IAA
|
Enhanced mineral uptake, increased diameter,
weight, length of ear, number of grains/rows, cob
weight and grain yield
|
|
Bradyrhizobium sp
|
Nitrogen fixation, siderophore
and IAA production, P
solubilisation
|
Increased vegetative parameters and seed yield in
mung bean
|
|
Rhizobium meliloti
|
Nitrogen fixation, production of
siderophore and chitinase
|
Increased growth and yield, pods quality and better
use of nitrogen in peanut
|
PHOSPHATE SOLUBILIZING PGPR
One of the most crucial nutrients for plants is phosphorus, which influences every biological function either directly or indirectly. P is essential for all major metabolic activities in plants, including respiration, energy transmission, photosynthesis, signal transduction, and molecular biosynthesis. Although soils contain a significant amount of P in both organic and inorganic forms, the availability of P is one of the primary factors limiting plant growth. This is because plants can absorb P from the soil solution as orthophosphate ions H2PO− 4 and HPO42−, but the majority of soil P is in insoluble state and is therefore unavailable (Soumare et al., 2020). Phosphorous biofertilizers include both phosphate solubilizers and mobilizers. Phosphate-solubilizing microorganisms (PSM) produce and release several chemicals that help solubilize inorganic P, such as tricalcium phosphate, hydroxyapatite, and rock phosphate. The release of organic acids, hydroxyl ions and CO2 are one method. This lowers the pH of the soil, directly dissolving the insoluble phosphates and causing PO42− to be exchanged for acid ions (Wei et al., 2018).Moreover, microbes could produce chelating substances, which can be used to release related soluble phosphates by capturing and mobilizing cations from various insoluble phosphates, such as Ca+2, Al+3, and Fe+3. PSM reduce the need for mineral P fertilizer inputs, which when used excessively can have detrimental effects on the environment, such as eutrophication of freshwater bodies, by raising P bioavailability. The most investigated P solubilizers belong to genera Pseudomonas, Bacillus, Rhizobium, Enterobacter, Penicillium, and Aspergillus (Anand et al., 2016; Riaz et al., 2020).
It has been reported that phosphorus in soil is solubilizedby a variety of microorganisms, such as Bacillus, Pseudomonas, Achromobacter, Brevibacterium, Erwinia sp., Flavobacterium sp., Micrococcus sp., Corynebacterium, Xanthomonas sp., Nostoc, Rhodococcus sp., Serratia phosphaticum, Acytonema, Calothrix brauna, and Tolypothrixceylonica, Bacillus, Pseudomonas, Achromobacter, Brevibacterium, Burkholderia, Sarcina sp., and Scytonema. Like this, it has been observed that a variety of fungi, such as Alternaria sp., Aspergillus sp., Cephalosporium sp., Fusarium sp., Rhodotrulaminuta, Saccharomyces cerevisiae, Torula thermophila, Paeciliomyces, Penicillium, and Sclerotium rolfisi, exhibit phosphate solubilization (Sindhu et al., 2014; Santoyo et al., 2021).
When chickpea was inoculated with PSB, the number of nodules per plant, shoot length, number of pods per plant, and grain production increased by 23%, 13%, 17%, and 15%, respectively. In addition, the stability of the soil aggregate increased by 37%, and 2.35 times more phosphorous was liberated from the rock phosphate. Additionally, the protein, phosphorus and nitrogen contents of chickpea and straw were increased to 16%, 42%, and 11%, respectively (Ditta et al., 2018).
In addition, mycorrhiza is essential for the cycling of nutrients, mobilization of phosphorus and the growth of microbial biomass. Native arbuscular mycorrhizae (AM) are primarily found in soil, where they colonize plant roots and promote plant growth. The availability of phosphorous increases abruptly when mycorrhiza is introduced into low-phosphorus soil. The primary function of arbuscular mycorrhizal fungi, which are found in nature in symbiotic relationships with cereals and horticultural plants, is to increase the bioavailability of phosphorus. Plants with mycorrhizal associations out compete those without in terms of utilizing soil nutrients because fungi have long enough hyphae to reach distant soils that plant roots are unable to access. Gigaspora, Acaulospora, Glomus, Scutellospora, and other genera are commonly used as biofertilizers (Pandey et al., 2019). Mycorrhiza has a beneficial effect on the structure of the soil as well. Improved plant health, soil productivity, soil aggregate stability and increased agricultural production are among the benefits of mycorrhizal inoculation.
POTASSIUM SOLUBILIZING PGPR
Potassium (K) is a key inorganic cation found in the cytoplasm of plants, and it is a crucial macronutrient for plants. It is involved in many basic metabolic processes, including protein synthesis, photosynthesis, and cell structure and function. In addition to being one of the most prevalent elements on Earth, potassium is the second most abundant nutrient in soil after nitrogen. Nonetheless, approximately 98% of soil K is found in a non-exchangeable state within the crystal structures of feldspar and mica (such as biotite and muscovite). Only 0.1–0.2% of the total is in the soil solution and immediately available for plant absorption, with the remaining 1-2% adsorbed onto organic matter and clay particles (Mitter et al., 2021). By solubilizing K, microorganisms can make it more accessible to plants, which is an important part of the K cycle. The release of organic acids, such as tartaric, citric, oxalic, gluconic, lactic, and malic acid, is the most well-known process of microbial K solubilization. (Macik et al., 2020).
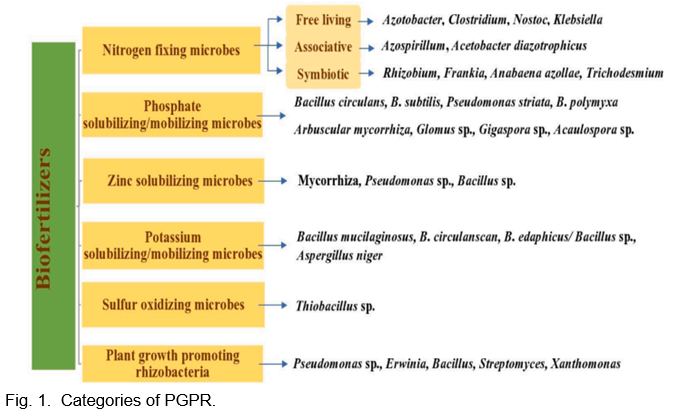
Due to their diversity and complexity in terms of phylogeny, microbes interact with plants through a variety of mechanisms, such as symbiosis, parasitism, amensalism and neutralism (Fig. 1). These microorganisms are collectively referred to as the plant microbiome because they are dependent on photosynthesis in plants and reciprocate by influencing plant growth. Since mycorrhizal fungi and PGPR have been proposed to be essential for both nutrient availability in the soil and alleviation of stress, these microorganisms have grown to be key components in the efficient operation of the soil ecosystem.
Certain phytohormones, or plant growth regulators, are synthesized by bacteria and plants in very small amounts. These hormones affect a variety of physiological processes, such as cell division, development, gene expression, and stress responses, as well as the shape, flowering, senescence, and seed growth. PGPR either produce hormones or change the hormone concentration in the plant in response to biotic and abiotic stressors. Many microbial species have been identified as producing gibberellins and cytokinin’s, which are bioactive hormones that influence a variety of developmental processes, including fruit setting, blooming, stem elongation, root hair growth, and seed germination (Khanet al., 2020). It was discovered that bacteria that produce siderophores that are essential for both biocontrol and growth stimulation. Siderophores are produced by Pseudomonas, Aeromonas, Azotobacter, Bacillus, Rhizobium, Azadirachta, Streptomyces, Burkholderia and Serratia.
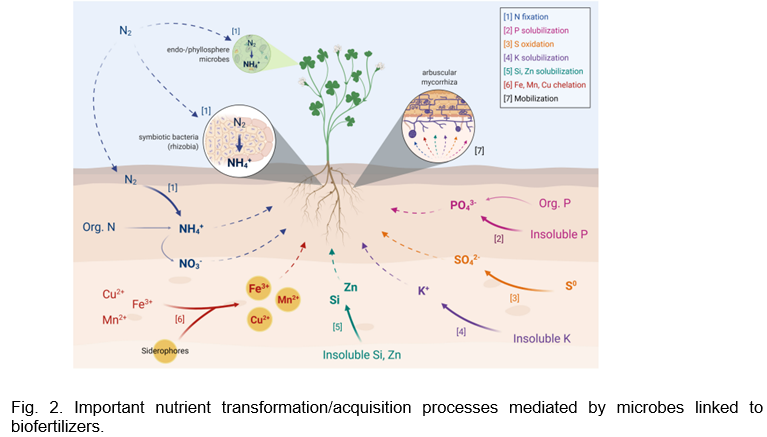
Arbuscular mycorrhizal fungi (AMF) are obligate symbionts which can create symbiotic associations with around 80% of land plant species, including crops used in agriculture. AMF provides various advantages, including improving the uptake of water and mineral nutrients (such as P, N, S, Cu, and Zn) by their host plants in return for a carbon source. The extraradical AMF mycelium expands the amount of soil that is explored by entering smaller soil holes in addition to extending well beyond the rhizosphere. This is especially critical for the uptake of less mobile nutrients like P, and AMF is widely recognized for its capacity to improve P uptake, particularly in soils lacking in P (Fig. 2.).
ADVANTAGES AND LIMITATIONS OF PGPR
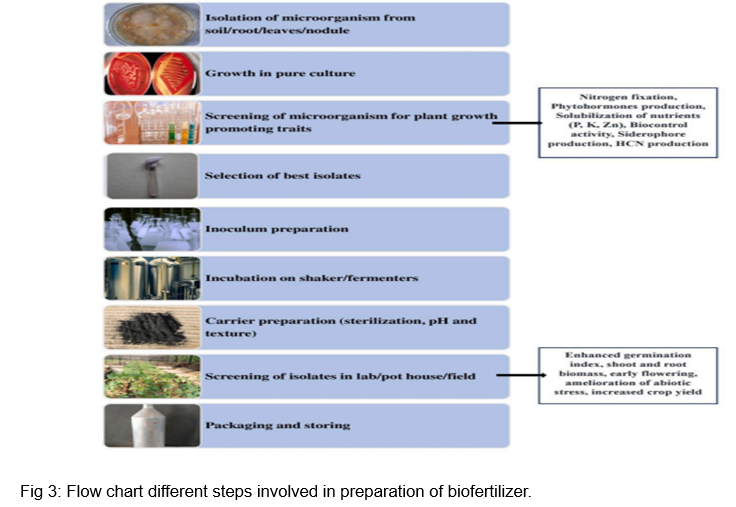
PGPR have the potential to significantly improve environmental sustainability by lowering soil, water, and air pollution brought on by chemical fertilizers. It improves soil fertility, boosts microbial biodiversity, aids in plant survival under a variety of stressful situations, and controls plant disease. Nevertheless, a few restrictions on PGPR prevented their widespread use and commercialization. The main obstacles to industrial-scale production and commercialization include the use of fertilizers with lower nutrient contents than chemical fertilizers, variations in or deviations from their efficacy when stored and exposed to external conditions such as light, temperature, and humidity, inappropriate carrier materials, inappropriate microbial strains, a lack of quality assurance, inexperienced and inefficient staff, low volume production, improper inoculation techniques, mutated strains, and market availabilityare the major limitations in the industrial scale production and commercialization of biofertilizers (Fig. 3).
CASE STUDIES
The experiment conducted during summer of 2018 at ICAR-Indian Agricultural Research Institute, New Delhi to study the effect of crop establishment practices and microbial inoculation on growth and nutrient uptake by summer green gram (Vigna radiata (L.) Wilczek). Results indicated that zero tillage with 2.5 t ha-1 residue produced significantly higher growth parameters like crop growth rate (CGR) and net assimilation rate (NAR) at 20, 35 and at 50 DAS, grain (968.6 kg ha-1) and (2415.3 kg ha-1) stover yield. The same treatment showed significantly higher uptake of N, P and K in grain and stover over other two treatments. Seed inoculation with Rhizobium+PSB+AM fungi was significantly superior to other two treatments about growth parameters like CGR and NAR at 20, 35 and 50 DAS, grain yield, stover yield and uptake of N, P and K in grain and stover of summer green gram. Highly positive correlations were recorded between grain yield and yield attributes like grain yield and pods/plant, grain yield and grains/ pod, grain yield and test weight of summer green gram (Naragund, et al., 2020).
Shome et al., 2022 conducted following a completely randomized design to explore the impact of nitrogen fixing bacteria and PSB on the growth, yield, and quality attributes of soybean alongside soil nutrient availability using Rhizobium japonicum and Pseudomonas striata. The experiment consisted of two factors, R. japonicum (100% N, R. japonicum alone or with 50% N and control) and P. striata (100% P, P. striata with 75% P and control). Results revealed a significant influence of interaction on seed N, yield, protein, oil, and nodules of soybean. Microbial inoculants with or without N and P fertilizers produced a statistically similar yield as 100% N and P. Furthermore, R. japonicum and P. striata along with 50% N and 75% P increased 7% protein and 19% oil than 100% N and P. R. japonicum enhanced soil N content and P. striata improved soil phosphorus availability. Overall, R. japonicum and P. striata inoculation with 50% N and 75% P can potentially improve the yield and the quality of soybean and soil nutrient conditions.
Shilpa et al., (2022) in her studies based on the utilization of carbon sources, thirty isolates were identified as Azospirillum brasiliense, and twenty isolates were identified as A. amazonense. Selected isolates were tested for their antifungal activity against Exserohilum turcicum causal agent of turcicum blight of maize under in vitro condition and antifungal activity of isolates was determined by the decreased diameter of fungal growth compared to the diameter of the control fungal growth. The isolate ACDAZS-35 showed the highest percentage inhibition of 85.56%. The isolate ACD-15 at different nitrogen levels showed a significant increase in number of leaves over the uninoculated control. It was found that maximum number of leaves, SPAD values, N content was observed in ACDAZS-9 inoculated with 50% N application at 30 and 60 DAS. The present study revealed that the selected Azospirillum inoculants could increase the growth of maize plants and can save 25% of inorganic nitrogen application.
Azolla has been exploited widely as biofertilizer for rice and some crop plants, and it fixes atmospheric nitrogen due to the presence of a symbiotic Cyanobacterium and Anabaena azollae. It has several other uses such as food, feed, biogas producer and hyper accumulator of heavy metals, etc. The use of biofertilizers like Azolla not only increased the crop productivity but also improved the long-term soil fertility. These multi-dimensional uses of Azolla-Anabaena system would be ideal and environment friendly in sustainable agriculture. The current study critically reviewed the Azolla’s potential capacity to be used in sustainable agriculture (Santhiya and Jeeva, 2022).
A study conducted by Reddy and Rahe (1989) monitored the populations of a marked strain of the B-2 isolate of B. subtilis (B-2) in the rhizospheres of onion seedlings grown from bacterized seeds in muck soil at various pH, moisture and temperature regimes for 14 weeks. Irrespective of regime, populations of B-2 in onion rhizospheres declined rapidly within the first 14 days after seeding, from 4.8×106 cfu per seed on day zero to a mean of 1.1×103 cfu per plant at day 14, and less rapidly during the next 12 weeks following seeding.
Various formulations are available in the market viz., carrier based, oil and liquid based formulations. This carrier must be nontoxic for plants and microbes, physically and chemically stable, cost-effective, biodegradable and able to maintain humidity, and ensure cell viability (Balla et al., 2022 and Mącik et al., 2020). Peat-based inoculants have historically dominated the commercial biofertilizer market owing to the substantial surface area of peat, its excellent water retention properties, and the conducive environment it provides for metabolic activity and cell proliferation during storage. Furthermore, due to their carbon dioxide trapping capacity, if peat is used as a carrier, its mitigation effect on climate change will be reversed when the captured CO2 is released again. In addition to their negative environmental impact on peat-rich ecosystems (John et al., 2011, Balla et al., 2022, Gewin, 2020). Thus, there has been a shift towards the development of new carriers from both organic materials (such as compost, biogas slurry, crushed corn cobs, biochar, peat, etc.) and inorganic substances (including zeolite, perlite, lignite, or vermiculite) (Thomas et al., 2019, John et al., 2011).
Field trials conducted over two growing seasons indicated that none of the 19 PGPR strains, applied as seed treatments at two concentrations, significantly reduced late leaf spot disease. The same chemical elicitors tested in the greenhouse, including BABA, did not elicit significant disease protection. Some combinations of four PGPR and BABA significantly reduced the disease at one but not at two sample times. Collectively, these results suggest that late leaf spot resistance in peanut is not systemically inducible in the same manner as is resistance to diseases in other crops by PGPR and chemical inducers Zhang et al., (2001).
A carotenoid producing halotolerant PGPR Dietzia natronolimnaea STR1 protecting wheat plants from salt stress by modulating the transcriptional machinery responsible for salinity tolerance in plants. The expression studies confirmed the involvement of ABA-signalling cascade, as TaABARE and TaOPR1 were upregulated in PGPR inoculated plants leading to induction of TaMYB and TaWRKY expression followed by stimulation of expression of a plethora of stress related genes. Enhanced expression of TaST, a salt stress-induced gene, associated with promoting salinity tolerance was observed in PGPR inoculated plants in comparison to uninoculated control plants. (Bharti et al., 2016).
ADVANCEMENTS IN PGPR
In the recent years, several advancements have developed to enhance the nutrients and growth dynamics of soil and plant for the betterment of crop production such as specialized microorganism are used for the inoculation of plants, best technologies are chosen, good carrier material for inoculum purposes. Furthermore, to reduce the cost of biofertilizer and for the sustainable production, current approaches are the artificial selection of microbiomes, mixed inoculants, PGPR as biofertilizers, use of biofilms as biofertilizers and Nano-biofertilizers.
MOLECULAR MECHANISM OF N2 FIXATION BY PGPR
Rhizobacteria is a nitrogen fixing bacteria that has developed symbiotic association and form nodules with the leguminous plants. They contain specialized genes known as Nif that carry out N2 fixation and are present in free existing as well as symbiotic bacteria. They also contain nitrogenase that includes reductase with Fe and Mo as cofactor and dinitrogenase reductase enzyme with iron cofactor. Dinitrogenase causes the reduction of nitrogen to ammonia by electrons that are facilitated by dinitrogenase reductase (Olanrewaju et al., 2017). Among Nif gene there are several regulatory transporters who contributed to the activation and formation of enzyme also regulate the synthesis of iron proteins. For the expression of Nif gene oxygen is an inhibitor but needed for the respiration of Rhizobium species. Oxygen causes the activation of the enzyme by binding with leg hemoglobin of bacteria with great affinity. Hence, to continue the process of nitrogen fixation accurately, significant amount of oxygen is facilitated to the rhizobacterium for respiration Basu et al., 2021). Genetic engineering is the easiest method to complete this process by transforming bacterial hemoglobin bounded with oxygen into bacteria. In result, respiratory rate of bacterial cells is increased up to several folds which in turn increase the rate nitrogen fixation to 68% (Gouda et al., 2018). Bacteria such as Azorhizobium and Rhizobium are symbiotic bacteria that fixes nitrogen by forming stem modules. They also enhance the growth of plant by synthesizing large amount of indole acetic acid. An efficient nitrogen fixer Bradyrhizobium when inoculated with the seeds of Mucuna, then it will enhance the potassium, Nitrogen, carbon content and phosphorus in the soil. Azotobacters are free living bacteria that produce different types of Phytohormones and vitamin B complex to promote pathogen resistance as well as growth of the crops (Sayyed et al., 2019). Azolla and Cyanobacteria are efficiently using as a biofertilizer because these species are useful as a source of nitrogen in the production of rice. Mainly Azolla is employed in the field before the cultivation of rice as a green fertilizer and supply essential amount of nutrients (Olanrewaju et al., 2017).
DEVELOPMENT OF MIXED PGPR
The use of mixture of bacterial strains instead of one is considered an important approach as different mechanism of action provide more potential for the use of biofertilizer. Mixed inoculant contains microbial consortia instead of single microbial strain that facilitate combined biofertilization effect and help plants to promote better uptake of nutrients. Recent studies have investigated a combined effect of co-inoculant containing Arbuscular mycorrhizal fungi (AMF) and rhizobacteria for the enhanced growth of the leguminous plant (Igiehon and Babalola, 2017). Co-inoculant of microbial specie allow wide range of biofertilizer efficiency and reliability for the fixation of nitrogen, phosphate solubilization and siderophore production and balanced growth and nutrition as compared to a single inoculant (Viswanath, 2019).
ENCAPSULATION AND LYOPHILISATION TECHNIQUE
Encapsulation technology is one of the current emerging techniques which allow the controlled release of biofertilizer into the environment. This technique is widely used to develop microbial species having regulating composition and structure characteristics (Schoebitz et al., 2013). Among this technology there is an advanced subfield of microencapsulation that works on the mechanism of constructing a protective covering such as capsule or a shell across an active molecule that could increase crop productivity and shelf life of biofertilizer. This type of formulation is more advantageous in term of slow and sustained release into soil and provide protection against environmental stress to biofertilizer (Çakmakçı, 2019).
Lyophilisation is the freeze-drying method for the preservation and storage of microorganism. It is widely used technique that helps to make a laboratory formulation of microorganisms. Lyophilisation is an efficient way to increase the survival rate of bacteria and storage for longer period; thus, the lyophilized microbial organisms can be used directly in combination with a suitable carrier in the field. This lyophilized structure can be used applied directly or a suitable carrier material can be used with it (Hernández et al., 2006).
NANO-PGPR
Nano-biofertilizer is the combined use of nano technology and biofertilizer for the more efficient and enhanced productivity of crops. The synergistic effect of nanomaterial and microbial fertilizer facilitates the maintenance of soil moisture and uptake of vital nutrients for plant (Kumari and Singh, 2019). Major issues of other fertilizers, such as instability on field due to fluctuation in environmental condition, temperature and change of pH, poor shelf life, reduced microbial strains, only short-term efficiency and need of large amount of fertilizer to cover wide area etc can lead to poor growth and provide less yield (Çakmakçı, 2019). The use of nano biofertilizer provide essential benefit to farmers in term of good field performance, low cost, less expenses, and provide more yield (Mala et al., 2017). Nano-biofertilizer are not only ecofriendly with the environment but also produce best quality crops by enhancing the efficiency of indigenous microbial population through utilization of essential nutrients such as potassium, nitrogen, and phosphorus. This will also stimulate the activity of microbial enzymes also contribute to the fertility of soil.
NANO-ENCAPSULATION TECHNOLOGY
Nano-encapsulation is the coating of biofertilizer at a nanoscale with the help of a nanomaterial. PGPR containing biofertilizers can be protected by using nanoencapsulation technology (Golbashy et al., 2017). This is a versatile technology which facilitates increased shelf life, provide controlled release of PGPR also allow controlled diffusion by biofertilizer. Nanomaterial is used for the nanoencapsulation of the nutrients and organic compounds such as zeolite, polymers and chitosan that facilitate the gradual and controlled bioavailability of nutrients to plants (Qureshi et al., 2018). Nano biofertilizer act synergistically with the microbial species or AMF inoculants through the enrichment of soil via bioavailability of nutrients that is carried out by several mechanisms such as fixation of nitrogen by enhanced activity of PGPR, phytohormone production, solubilization of phosphorus, and production of chelating agents that can chelate the elements and release them for plant uptake (Mala et al., 2017).
BIOFILM PGPR
Biofilms are the microbial communities that are be either biotic or abiotic and embed with some biological component that facilitates sustainability and structural characteristic to the biofilm (Junaid and Khan, 2018). Microbial species work together by making linkage with each other through the process of quorum sensing. The main objective for the development of biofilm biofertilizer is to overcome the problems of biotic and a biotic component of soil and to make the indigenous microbial population more resistive against pathogens and inhibitors (Parween et al., 2017). Biofilm is either synthesized on the plant surface or inside the plant and becomes advantageous for the fixation of nitrogen by PGPR and enhance the productivity of crops. Biofilm also help in Zn solubilization and production of chelating agents (Igiehon and Babalola, 2017). Biofilms provide a way to transfer the genetic material and organic nutrients between microbial species in a community. As compared to a biofilm comprising of single microbial species, multiple microbial strains in a biofilm are investigated to be more resistant and sustainable approach. For instance, the uptake of nutrient and tolerance to environmental stress is significantly increased for bacterial-fungal biofilm as compared to those microorganisms that are single or do not form biofilm (Velmourougane et al., 2017). Biofilm biofertilizer provide resistance to crops against heavy metal, drought, stress, pH, temperature, and attack of pathogen. Biofilm biofertilizer also help in the preservation of land and lessen harmful consequences on the human health. In addition to this, it also increases the productivity of the crops under different environmental conditions and facilitates longer shelf life to farmers (Mitter et al., 2021).
PGPR MARKET SIZE AND TRENDS
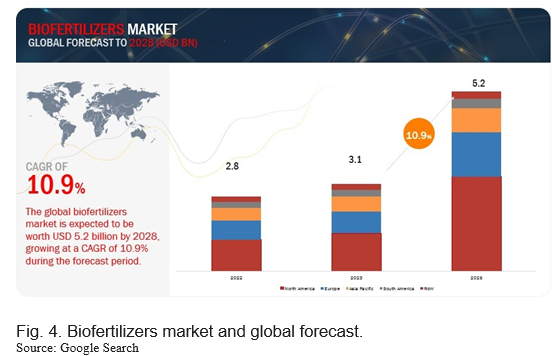
The global PGPR market was valued at 2.8 billion in 2022 and is projected to reach USD 5.2 billion by 2028, growing at a CAGR of 10.9% during the study period. The market size is estimated to be valued at USD 3.1 billion in 2023 (Fig. 4). Through natural processes such as nitrogen fixation; phosphate solubilization; and the production of compounds that stimulate growth, PGPR provides nutrients to plants. They boost soil organic matter and aid in restoring the soil's natural nutrition cycle. By using PGPR, growers can increase the sustainability and health of the soil and cultivate healthy crops. Only bacteria and other biological elements that are not damaging to the environment are present in PGPR. As a result, they contribute to reducing pollution brought on by agricultural activities, particularly soil pollution.
SMART PGPR IN SUSTAINABLE AGRICULTURE
The first Green Revolution (1965–1985) and the post Green Revolution period (1985≈2005) resulted in tremendous yield growth in staple crops, helping to feed the world despite large population increases and decreasing land availability for agriculture. The classical breeding methods and advancements in genomics, genome sequencing, transgenics, genome editing, and related breeding methodologies such as marker-assisted selection and incorporation of phenomics, data analytics, and artificial intelligence for the rapid development of climate-resilient crops. The utilisation of newer methods to understand essential genetic and molecular components of the multiple plant microbe interactions (PM) is required for their application. The use of clustered regularly interspaced short palindromic repeats (CRISPR)/Cas-mediated genome editing (GE) techniques to investigate PM interactions is of tremendous interest. The implementation of GE techniques to boost the ability of microorganisms or plants for agronomic trait development will be enabled by a comprehensive understanding of PM interactions. As a result, utilising the plant microbiota is a viable strategy for the next Secondary Green Revolution for meeting food demand.
MODE OF ACTION OF PGPR
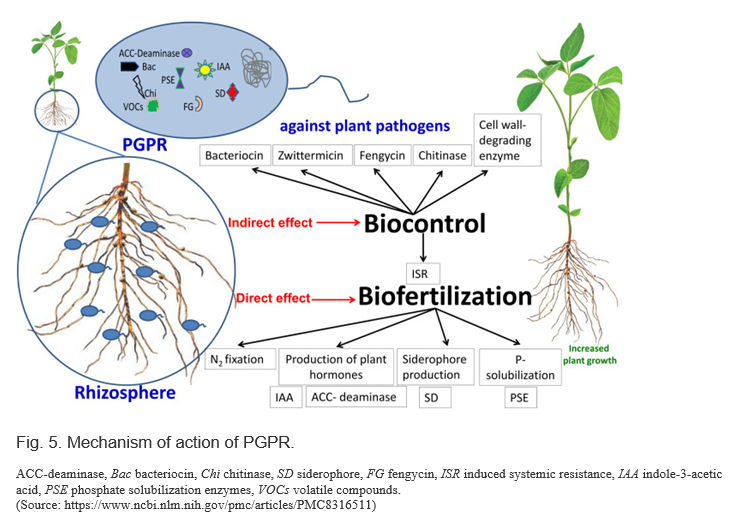
The mode of action of PGPRs on crop plants can be direct or indirect. Direct mechanisms include biofertilization (nitrogen (N2) fixation, production of plant hormones and siderophores (SDs), phosphorus (P) solubilization, etc.), root growth stimulation, rhizoremediation, and the control of plant stress. Additionally, PGPRs produce metabolites, such as indole-3-acetic acid (IAA), 1-aminocyclopropane-1-carboxylate-deaminase (ACC-deaminase), phosphate solubilizing enzyme (PSE), and SDs. They can also indirectly enhance plant growth by reducing the detrimental effects of phytopathogens through induced systemic resistance (ISR) and the production of antimicrobial compounds, such as bacteriocin, zwittermicin, fengycin, chitinase, and cell wall-degrading enzymes (Fig. 5)
FORMULATIONS OF PGPR
From many years in the field of agriculture application, biofertilizers has made higher achievements for the sustainable production of crops and establish more resilient conditions for the growth of microbes. But the use of conventional fertilizer formulation led to several pathogenic problems and reduced viability of microbial species during the field application (Çakmakçı, 2019). Therefore, the advanced suitable formulation for making a bioinoculant is essential so that the beneficial indigenous soil population can effectively develop under plant roots. For that purpose, microorganism need such type of protective formulation strategies that are effective at the target site and provide benefit to farmers (Stamenković et al., 2018). Formulation needs to be stable for longer period and should remain viable during the process of synthesis, packaging, transport and on field application (Mitter et al., 2021).
There are different PGPR formulations related to its recent approaches such as metabolite formulations, liquid formulations, solid carrier-based formulations, synthetic polymer-based formulations, granular formulations, powered formulations. The bioinoculant formulation is the uniform mixture of selected beneficial strain with a suitable carrier that can provide stabilization and protection of strain during transport and storage. The carrier is the vehicle of living latent microbes that provides protection and supportive niche to the microbial community. A good bioformulation should be effective, non-polluting, readily biodegradable with high water retention capacity and sufficient shelf life. There are still lots of potential of using innovative technologies towards improving the capability of bioinoculant, but it needs better integration of ideas and utilizing them for sustainable agriculture.
REGULATORY CONSTRAINS OF PGPR
Although the PGPR technology is a low cost, eco-friendly technology, several constraints limit the application or implementation of the technology. The constraints may be environmental, technological, infrastructural, financial, human resources, unawareness, quality, marketing, etc. The different constraints, in one way or another, affect the technique at production or marketing or usage. The guideline limits itself to only biofertilizers or biofertilizers containing carrier based (solid or liquid) living microorganisms which colonize the rhizosphere or the interior of the plant and promotes growth by increasing the supply or availability of primary nutrient and/or growth stimulus to the target crop, when applied to seed, plant surfaces, or soil. Throughout the guidelines of many countries, biofertilizers will refer to microbial biofertilizers. Common data requirements and equivalent registration systems pave the way for harmonisation of biofertilizer registration either within a region or across the entire continent. Flexibility is provided in this framework for registration upon the extent of regional integration. Our understanding is that we do not need any registration for PGPR based biofertilizers. Our Asian PGPR Society for Sustainable Agriculture (www.asianpgpr.com) is being developing guidelines for registration of PGPR as biofertilizers around the globe.
SPECIFIC RESEARCH NEEDS ON PGPR
1. Selection of effective and competitive multi-functional PGPR for a variety of crops.
2. Quality control system to produce PGPR and their application in the field, to ensure and explore the benefits of plant-microorganism symbiosis.
3. Study of microbial persistence of PGPR in soil environments under stressful conditions
4. Agronomic, soil, and economic evaluation of PGPR for diverse agricultural production systems.
5. Transferring technological know-how on PGPR production to the industrial level and for optimum formulation.
6. Establishment of "Bio-fertilizer Act" and strict regulation for quality control in markets and application.
CONCLUSIONS
PGPR offer a sustainable alternative, harnessing microbial power to enhance soil fertility and plant growth without adverse effects. Understanding soil ecology and microbial dynamics is crucial for optimizing PGPR efficacy. Leveraging genetic and biotechnological advancements can lead to improved formulations and production methods, paving the way for start-ups to explore this promising field. The application of inorganic chemical fertilizers was thus significantly reduced to 30-50%. This helps in the realization of environmentally friendly and sustainable agriculture.
Excess nutrients are accumulated in soils, particularly P because of over application of chemical fertilizers by farmers during intensive agricultural practices. Hence, major research focus should be on the production of efficient and sustainable bio-fertilizers for crop plants, wherein inorganic fertilizer application can be reduced significantly to avoid further pollution problems. In view of overcoming this bottleneck, it will be necessary to undertake short-term, medium, and long-term research, in which soil microbiologists, agronomists, plant breeders, plant pathologists, and even nutritionists and economists must work together.
Further research on the nano-encapsulation, multiple biofilms formation and mixed microbial PGPR are continuing to engineer disease resistance indigenous microorganism and plants. Future goal of PGPR is to develop innovative and eco-friendly agricultural techniques that are cheaper than use of chemical fertilizers. Thanks to the recent advances in molecular systems PGPR and the development of new-generation technology platforms for high efficacy microbial ecology studies, a collaborative, cross-disciplinary research effort, from ecological to bioprocess and metabolic engineering, will pave the way for predictable microbial diversity manipulation and new microbial-driven solutions to tackle the current environmental challenges we are facing at the nexus of food, water and energy.
REFERENCES
Anand, K. U., B. A. Kumari and M.A. Mallick. 2016. Phosphate solubilizing microbes: an effective and alternative approach as biofertilizers. Int. J.Pharm. Pharm. Sci. 8(2): 37-40.
Asoegwu, C. R, C. G. Awuchi, K. C. Nelson, C. G. Orji, O. U. Nwosu and U. C. Egbufor. 2020. A review on the role of biofertilizers in reducing soil pollution and increasing soil nutrients. Himalayan Journal of Agriculture. 1(1): 34-8.
Adeleke, R. A., A. R. Raimi, A. Roopnarain and S. M. Mokubedi. 2019. Status and prospects of bacterial inoculants for sustainable management of agroecosystems. p. 137-72. In B. Giri, R. Prasad, Q. S. Wu and A. Varma (eds.), Biofertilizers Sustain. Agric. Environ. Soil Biol. Springer, Cham, 10.1007/978-3-030-18933-47.
Bala, A., N. Karanja, M. Murwira, L. Lwimbi, R. Abaidoo and K. Giller. 2011. Production and use of Rhizobial inoculants in Africa, Www.N2 Africa.Org. 1–22.
Balla, A., A. Silini, H. Cherif-Silini, A. Chenari Bouket, F. N. Alenezi and L. Belbahri. 2022. Recent Advances in Encapsulation Techniques of Plant Growth-Promoting Microorganisms and Their Prospects in the Sustainable Agriculture. Appl. Sci. 12 (18), 9020. https://doi.org/10.3390/app12189020.
Basu, A., P. Prasad, S. N. Das, S. Kalam, R. Z. Sayyed, M. S. Reddy and E.l.H. Enshasy. 2021. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: recent developments, constraints, and prospects. Sustainability. 13(3): 1140. https://doi.org/10.3390/su13031140.
Bharti, N., S. S. Pandey, B. Deepti, V. K. Patel and A. Kalra. 2016. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci Rep. 6: 34768. https://doi.org/10.1038/srep34768.
Bourion, V., K. Heulin-Gotty, V. Aubert, P. Tisseyre, M. Chabert-Martinello, M. Pervent, C. Delaitre, D. Vile, M. Siol, G. Duc and B. Brunel 2018. Co-inoculation of a pea core-collection with diverse rhizobial strains shows competitiveness for nodulation and efficiency of nitrogen fixation are distinct traits in the interaction. Frontiers in Plant Science. 8: 290940. doi: 10.3389/fpls.2017.02249.
Çakmakçı, R. 2019. A Review of Biological Fertilizers Current use, New Approaches, and Future Perspectives. International Journal of Innovative Studies in Sciences and Engineering Technology. 5(7): 83-92
Chansa-ngavej, C. and S. Assavavipapan. 2007. Forecasting of rhizobial biofertilizer technology using maturity mapping. Proceedings of the Int. Colloq. Bus. Manag., Bangkok, Academia. P. 1-32
Ditta, A., M. Imtiaz, S. Mehmood, M. S. Rizwan, F. Mubeen, O. Aziz, Z. Qian, R. Ijaz and S. Tu. 2018. Rock phosphate-enriched organic fertilizer with phosphate-solubilizing microorganisms improves nodulation, growth, and yield of legumes. Communications in Soil Science and Plant Analysis. 49(21): 2715-25.
Garcha, S. and P. K. Maan. 2017. Biological nitrogen fixation in cereals crops: a bacterial perspective. Advances in Soil Microbiology: Recent Trends and Future Prospects: Soil Microbe Plant Interaction. 6(2): 127-51.
Graham, P. H., and C. P. Vance. 2003. Update on legume utilization legumes: importance and constraints to greater use. Plant Physiol. 131. p. 872-877, 10.1104/pp.017004.872.
García-Fraile, P., E. Menéndez and R. Rivas. 2015. Role of bacterial biofertilizers in agriculture and forestry. AIMS Bioeng. 2. p. 183-205, 10.3934/bioeng.2015.3.183.
Gewin, V. 2020. How peat could protect the planet. Nature. 578: 204–208.
Gouda, S., R. G. Kerry, G. Das, S. Paramithiotis, H. S. Shin and J. K. Patra. 2018. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiological Research, 206: 131–140. https://doi.org/10.1016/j.micres.2017.08.016.
Hernández, A, F. Weekers, J. Mena, C. Borroto and P. Thonart. 2006. Freeze-drying of the biocontrol agent Tsukamurella paurometabola C-924 predicted stability of formulated powders. Industrial Biotechnology. 2(3): 209-212.
Hegde, G. M., S. Aditya, D. Wangdi and B. K. Chetri. 2022. Mycoremediation: A Natural Solution for Unnatural Problems. In: V. R. Rajpal, I. Singh and S. S. Navi, S.S. (eds) Fungal diversity, ecology and control management. Fungal Biology. Springer, Singapore. https://doi.org/10.1007/978-981-16-8877-517.
Igiehon, N. O. and O. O. Babalola. 2017. Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Applied Microbiology and Biotechnology. 101(12): 4871–4881. https://doi.org/10.1007/s00253-017-8344-z.
Jain, D., J. Sharma, G. Kaur, A. A. Bhojiya, S. Chauhan, V. Sharma, A. Suman, S. R. Mohanty and E. Maharjan. 2021. Phenetic and molecular diversity of nitrogen fixating plant growth promoting Azotobacter isolated from semiarid regions of India. Bio.Med. Research International. 5(3): 221-29.
John, R. P., R. D. Tyagi, S. K. Brar, R. Y. Surampalli and D. Prévost. 2011. Bio-encapsulation of microbial cells for targeted agricultural delivery. Crit. Rev. Biotechnol. 31: 211-226.
Junaid, P. M. and F. Khan. 2018). Biofilm: A future generation biofertilizer. Indian Journal of Applied and Pure Biology. 33(2): 131-134.
Khan, N., A. Bano, S. Ali and M. A. Babar. 2020. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regulation. 90: 189-203.
Kumari, R., and D. P. Singh. 2019. Nano-biofertilizer: An emerging eco-friendly approach for sustainable agriculture. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences. 1-9.
Mala, R., R. Celsia Arul Selvaraj, V. Barathi Sundaram, R. Blessina Siva Shanmuga Rajan and U. Maheswari Gurusamy. 2017. Evaluation of nano structured slow-release fertilizer on the soil fertility, yield and nutritional profile of vigna radiata. Recent Patents on Nanotechnology. 11(1): 50-62.
Lawal, T. E. and O. O. Babalola. 2014. Relevance of biofertilizers to agriculture J. Hum. Ecol. 47: 35-43, 10.1080/09709274.2014.11906737.
Macik, M., A. Gryta and M. Frac. 2020. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Advances in Agronomy. 16 (2): 31-87.
Mfilinge, A., K. Mtei and P. Ndakidemi. 2014. Effect of Rhizobium inoculation and supplementation with phosphorus and potassium on growth and total leaf chlorophyll (Chl) content of bush bean Phaseolus vulgaris, L. Agricultural Sciences. 5: 1413-1426
Effect of Rhizobium Inoculation and Supplementa-
tion with Phosphorus and Potassium on Growth and Total Leaf Chlorophyll (Chl) Content of Bush Bean Phaseolus vulgaris, L.
Mishra, P. and D. Dash. 2014. Rejuvenation of biofertilizer for sustainable agriculture and economic development. Consilience. 1(11): 41-61.
Mitter, E. K., M. Tosi, D. Obregón, K. E. Dunfield and J. J. Germida. 2021. Rethinking crop nutrition in times of modern microbiology: innovative biofertilizer technologies. Frontiers in Sustainable Food Systems. 5: https://doi.org/10.3389/fsufs.2021.606815.
Malusà, E., L. J. Sas-Paszt and Ciesielska. 2012. Technologies for beneficial microorganisms inocula used as biofertilizers Sci. World J. 1-12: 10.1100/2012/491206.
Naragund, R., Y. V. Singh, R. S. Bana, A. K. Choudhary, J. Pranita, K. Praveen, and S. Andhimansu. 2020. Influence of crop establishment practices and microbial inoculants on plant growth and nutrient uptake of summer green gram (Vigna radiata) Annals of Plant and Soil Research. 22(1): 55-59.
Olanrewaju, O. S., B. R. Glick and O. O. Babalola. 2017. Mechanisms of action of plant growth promoting bacteria. World Journal of Microbiology and Biotechnology. 33(11): 197. https://doi.org/10.1007/s11274-017-2364-9.
Pandey, D., H. K. Kehri, I. Zoomi O. Akhtar and A. K. Singh. 2019. Mycorrhizal fungi: Biodiversity, ecological significance, and industrial applications. Recent Advancement in White Biotechnology Through Fungi: Diversity and Enzymes Perspectives. 1: 181-99.
Pereg L, L. E. de-Bashan and Y. Bashan. 2016. Assessment of affinity and specificity of Azospirillum for plants. Plant and Soil. 39(9): 389-414.
Riaz, U., S. M. Mehdi, S. Iqbal, H. I. Khalid, A. A. Qadir, W. Anum, M. Ahmad and G. Murtaza. 2020. Bio-fertilizers: eco-friendly approach for plant and soil environment. Romberger, J. A. and P. Mikola. 1966. International Review of Forestry Research. Academic Press, New York. 10.2307/forhis/10.2.20-a.
Reddy, M. S. and H. Singh. 2011. Plant growth-promoting rhizobacteria (pgpr) for sustainable agriculture. Field evaluation of P solubilising bacteria on the growth and yield of wheat and maize grown in rock phosphate amended soils - Proceedings of the 2nd Asian PGPR Conference August 21-24. Beijing, P. R. China.
Reddy, M. S. and J. E. Rahe. 1989. Growth effects associated with seed bacterization not correlated with populations of Bacillus subtilis inoculant in onion seedling rhizospheres. Soil Biology and Biochemistry. 21(3): 373-378.
Strijdom, B.W. 1998. South African studies on biological nitrogen-fixing systems and the exploitation of the nodule bacterium-legume symbiosis. S. Afr. J. Sci. 94: 11-23.
Santoyo, G., P. Guzmán-Guzmán, F. I. Parra-Cota, S. D. Santos-Villalobos, M. D. Orozco-Mosqueda and B. R. Glick. 2021, Plant growth stimulation by microbial consortia. Agronomy. 11(2): 219. https://doi.org/10.3390/agronomy11020219.
Sindhu, S. S., M. Phour, S. R. Choudhary and D. Chaudhary. 2014. Phosphorus cycling: prospects of using rhizosphere microorganisms for improving phosphorus nutrition of plants. In: Parmar, N., Singh, A. (eds). P. 199-237. Geomicrobiology and Biogeochemistry. Soil Biology, vol 39. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-41837-2_11 199-237.
Soumare, A., K. Boubekri, K. Lyamlouli, M. Hafidi, Y. Ouhdouch and L. Kouisni. 2020. From isolation of phosphate solubilizing microbes to their formulation and use as biofertilizers: status and needs. Frontiers in bioengineering and biotechnology. 7: 425-35.
Sanhthosh, K. R. G. and M. Sandeep. 2020. Biofertilizers role in major pulse crops of India International Journal of All Research Education and Scientific Methods (IJARESM), ISSN: 2455-6211 Volume 8, Issue 11.
Shome, S., A. Barman and Zakaria M. Solaiman. 2022. Rhizobium and Phosphate Solubilizing Bacteria Influence the Soil Nutrient Availability, Growth, Yield, and Quality of Soybean (Bangaldesh) Agriculture. 12: 1136. https://doi.org/10.3390/ agriculture 12081136.
Santhiya, B. and S. Jeeva. 2022. Azolla as a source of biofertilizer for sustainable crop production. Journal of Xi’an Shiyou University, Natural Science Edition ISSN: v1673-064X. 18 (11) 598-605.
Schoebitz, M., M. D. López and A. Roldán. 2013. Bioencapsulation of microbial inoculants for better soil–plant fertilization. A review. Agronomy for Sustainable Development. 33(4): 751-765.
Sayyed, R. Z., M. S. Reddy and S. Antonius. (Eds.). 2019. Plant growth promoting rhizobacteria (PGPR): Prospects for Sustainable Agriculture. Springer Singapore.
Shilpa, H., V. Navi, G. P. Santhosh and G. M. Hegde. 2022. Isolation, identification of Azospirillum and its inoculation effects on maize (Zea mays L.) J. Farm Sci. 35(3): 370-380.
Stamenković, S., V. Beškoski, I. Karabegović, M. Lazić and N. Nikolić. 2018. Microbial fertilizers: A comprehensive review of current findings and future perspectives. Spanish Journal of Agricultural Research. 16(1), e09R01. https://doi.org/10.5424/sjar/2018161-12117.
Thomas, L. and I. Singh. 2019. Microbial biofertilizers: types and applications. pp. 109-135. In Giri, B., R. Prasad, Q. Wu and A. Varma (Eds.), Biofertilizers Sustain. Agric. Environ., Vol 55, Springer Nature, Switzerland AG. 10.1007/978-3-030-18933-41.
Viswanath, B. 2019. Recent Developments in Applied Microbiology and Biochemistry. p.363-373.
Velmourougane, K., R. Prasanna and A. K. Saxena. 2017. Agriculturally important microbial biofilms: present status and prospects. Journal of Basic Microbiology: 57(7): 548-573.
Wei, Y., Y. Zhao, M. Shi, Z. Cao, Q. Lu, T. Yang, Y. Fan and Z. Wei. 2018. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresource Technology. 24 (7):190-99.
Zhang, S., M. S. Reddy, N. Kokalis-Burelle, L. W. Wells, S. P. Nightengale and J. W. Kloepper. 2001. Lack of induced systemic resistance in peanut to late leaf spot disease by plant growth-promoting rhizobacteria and chemical elicitors. Plant Dis. 85: 879-884.
Recent Advances in Biofertilizers and Their Application in Sustainable Agriculture
This manuscript was originally presented at the NACF, MARDI, IFOAM-Asia, and FFTC workshop titled “Technology, Implementation, and Policy Review of Biofertilizers and Biopesticides in the Asian and Pacific Region,” held at the NACF Headquarters in Seoul, Korea, on May 8–10, 2024.
ABSTRACT
With the increase in world population, the demography of humans is estimated to be exceeded, and it has become a major challenge to provide an adequate amount of healthy food globally. The use of chemical fertilizers and fungicides causes the plant to grow efficiently and rapidly to meet the food demand. But the drawbacks of using a higher quantity of chemicals are environmental pollution, changes in the soil ecology, physiochemical composition, decreasing agricultural yields leading to several health hazards. Climatic factors such as abiotic and biotic stresses like soil salinity, drought, wind, raising temperatures, heavy metals, waterlogging, and different weeds and phytopathogens like bacteria, viruses, fungi, and nematodes which attack plants, reducing crop productivity and quality. In recent years, the growing adoption of organic farming has resulted in a more sustainable approach towards agriculture as it emphasises on adopting environmentally friendly practices to ensure food security while protecting and nurturing soil health. Various strategies to make organic and sustainable agriculture, one of the key factors being the use of biofertilizers (PGPR) to provide optimum nutrients to crop and enhance soil health. There is a shift toward the use of biofertilizers due to all these facts, which provide nutrition through natural processes like zinc, potassium and phosphorus solubilization, nitrogen fixation, production of hormones, siderophore, various hydrolytic enzymes and protect the plant from different plant pathogens and stress conditions. They are made up of physiologically active microbes, such as bacteria, algae, and fungal species, that help to improve, preserve, and transform nutrients from an unusable to a useful form. There are various ways to introduce biofertilizers, such as root dipping, in-furrow soil application and inoculating onto seeds with a liquid or dry formulation. The recently emerging techniques such as nano-biofertilizer and biofilm biofertilizer enable crops to become more resistant toward environmental inhibitors and pathogenic components. The advanced microbial inoculants by formulating various plant growth-promoting rhizobacteria (PGPR) to combat plant diseases and environmental stress is getting more popular in recent days. They are often regarded as a renewable source of plant nutrients and promote soil health and have become a vital constituent in an integrated crop and soil management systems. The demand for biofertilizers in Asia-Pacific is anticipated to increase throughout the forecast period, with a CAGR of 8.6%. This review critically examines the current state-of-art on use of PGPR’s in different crop in maintaining soil fertility and enhancing crop productivity. It is believed that acquisition of advanced knowledge of plant-PGPR interactions, bioengineering of microbial communities to improve the performance of PGPR under field conditions, will help in devising strategies for sustainable, environment-friendly and climate smart agricultural technologies to deliver short and long terms solutions for improving crop productivity to feed the world in a more sustainable manner.
Keywords: Biofertilizers, Sustainable crop production, Bioactive hormones, Plant growth promotion, Microbial inoculants, PGPR
INTRODUCTION
In a world where global food security is increasingly threatened, adopting sustainable and environmentally friendly Bio-Agri Inputs approaches to safeguard our agricultural crops is of paramount importance. Addressing these pressing issues necessitates a paradigm shift in agricultural practices, promotion of environmentally friendly alternatives for the reduction of chemical inputs is a mandatory approach. In this context, leveraging the interactions between plants and beneficial microorganisms, such as plant growth-promoting rhizobacteria (PGPR), has emerged as a promising strategy and in this context, we refer them as biofertilizers. PGPR, a diverse group of soil-dwelling bacteria, are renowned for their multifaceted roles in enhancing plant growth, improving nutrient uptake, and conferring resistance to various biotic and abiotic stresses. They have the potential to serve as biofertilizers and or biocontrol agents which are substitutes for undesirable chemicals and pesticides, promoting sustainable healthy practices for sustainable and organic agriculture. Over the years, the research has uncovered an intriguing facet of the plant-PGPR interaction, namely, the phenomenon of plant defence priming. Biofertilizers are biologically active formulations comprising beneficial live microbes (PGPR) in economical carrier materials, which when applied to the soil, plant, and seed improve plant growth and development by enhancing the supply of plant growth-promoting substances or nutrients. The term “biofertilizer” has been defined in different ways during the past 20 years, which derives from the improved understanding of the relationships occurring between the rhizosphere microorganisms and the plant. Finally, there is a common consensus among many researchers include ours is that “a substance which contains living microorganisms which, when applied to seed, plant surfaces, or soil, colonizes the rhizosphere or the interior of the plant and promotes growth by increasing the supply or availability of primary nutrients to the host plant”.
Plant biostimulants are becoming popular in the agricultural market as products to increase growth, health, and/or yield of crop plants. The challenge is that there are many types of biostimulants, each with unique mechanisms of action, and these dynamics make it difficult to optimize product placement into grower systems. A global assessment of the structure and function of the crop microbiome is urgently needed for the development of effective and rationally designed microbiome technologies for sustainable agriculture. Such an effort will provide new knowledge on the key ecological and evolutionary interactions between plant species and their microbiomes that can be harnessed for increasing agriculture productivity. Hower, in this review, whatever the term may be, we will be focused on PGPR as a biofertilizer which includes as bioinoculants, biostimulants, biofilm and plant microbiome for ease of understanding.
The groups of microbes mostly used for inoculant production are the arbuscular mycorrhizal (AM) fungi, PGPR and nitrogen-fixing rhizobia (Malusà, et al., 2012). Most soils contain extremely elevated levels of bacteria (often estimated to be ~108–109 bacterial cells per gram of soil). Some of these bacteria are PGPB, some are phytopathogens and some do not have any discernible effect on plant growth. The knowledge of the physiological traits and mechanisms that are used by PGPR to promote plant growth has developed over the past 20–30 years. In fact, while only a small number of PGPR have been commercialized as adjuncts to agricultural practice, both the number of commercialized strains and the amount of farmland devoted to using this approach continue to increase with each succeeding year.
All land plants host a microbiome composed of bacteria, fungi, oomycetes, viruses, archaea, and protists. These organisms primarily colonize the root environment, especially the rhizosphere (the soil portion immediately adjacent to the root), the rhizoplane (intended as the root surface), and to a lesser extent the leaf surface (phyllosphere), seed (spermosphere), and internal (endosphere) plant environments. Therefore, microbiomes associated with plants show a different degree of intimacy with the plant: while epiphytic microorganisms can live externally, endophytic microorganisms efficiently colonize the internal plant tissues. In this context, plants may be viewed as superorganisms that include their microbiomes, which provide several unique functions and features. However, notwithstanding an abundance of recent studies, the knowledge on plant-associated microbiomes and their detailed effects on crop productivity, growth, health, and disease are still scanty. Plants can take advantage of the rhizosphere microbiome by selectively inducing activities within microorganisms that support their development and survival. In this way, rhizodeposition shapes the plant-associated microbiome, which in turn, can affect both plant metabolic pathways and exudate release. Based on this knowledge, the existence of crosstalk among the host–microbiome and metabolome, where plants perceive and understand the chemical communication among microbes and vice-versa. By considering the plants as an holobiont communicating with the external environment, that in turn affects the interrelationships occurring in the holobiont itself, where molecules affecting rhizosphere interactions and plant health can be degraded, used, and modified by the members of the rhizosphere microbiome.
Until now, most of the knowledge and understanding of how PGPR promote plant growth has included studies of isolated bacteria examined in controlled laboratory conditions that have not considered the range of organisms and abiotic conditions that affect how these bacteria function in soil. In the soil, PGPR typically acts together in groups or consortia, with the consortia being responsible for facilitating plant growth, that is, different bacteria within the consortia satisfy different plant needs. The attraction of both individual PGPR, as well as bacterial consortia of PGPR, to a particular plant is dictated, in part, by the range and concentration of small molecule root exudates produced by a particular plant. Interestingly, even though the microbiota in most microbiomes contains both PGPR and deleterious microorganisms plants attract beneficial PGPR and not phytopathogens. Thus, despite the presence of phytopathogens in many microbiomes, the beneficial PGPR often protects plants from the deleterious effects of phytopathogens.
The world's population is continually growing, and projections indicate that by 2050, there will be approximately 9.7 billion people on the planet. This quick expansion is closely linked to extensive urbanization, industrialization, and agricultural output (Macik et al., 2020). Numerous hybrid crop varieties have been developed and are still being developed to meet the demands of the expanding population.
The efficient application of chemical fertilizers to crops yielding undeniable benefits for crop growth, yield, and quality as well as financial gain for farmers. Unfortunately, because of the greater quantities of synthetic fertilizers are being used, leads to soil, water, and air pollution which could be a serious threat to the ecosystem. These synthetic chemicals are being applied carelessly causes their accumulation below ground. This leads to adverse changes in the structure, fertility, and water-holding capacity of the soil. In-addition, overuse of synthetic fertilizers is also strongly associated with the greenhouse effect and eutrophication of water bodies. Extended usage of mineral fertilizers can cause the soil's nutrient level to decrease and increase crop susceptibility to several diseases (Macik et al., 2020).
For better yields, farmers applying lot of chemical fertilizers that can increase yield but damage the soil strength and biota. To overcoming these problems recent agricultural practices mainly utilization of biofertilizers such as Rhizobium strains, PSB (phosphate solubilizing bacteria), VAM (Vascular Arbuscular Mycorrhiza), Cyanobacteria and various other microorganism plays a vital role on crops in increasing nutrients uptake by fixation of nutrients by forming nodules in roots that helps in mineralization process that can convert the unavailable form of nutrients to available form that can easily uptake by plant. These biofertilizers are mostly host specific that can symbiont with pulse crop and boost the growth and yield attributes (Santhosh and Sandeep, 2020).
TheGeenera Gigaspora, Glomus, Bacillus, Pseudomonas, Azotobacter, Azospirillum, Bradyrhizobium and Rhizobium, are commonly used as PGPR (Adeleke, 2019, Thomas and Singh, 2019). These PGPR play essential roles in soil ecosystem functions such as restoring and preserving soil nutrient richness. Several studies have been conducted on PGPR and their various benefits in sustainable agriculture, especially in promoting plant nutrition (García-Fraile, 2015, Lawal and Babalola, 2014). A more environmentally friendly option to chemical fertilizers is PGPR or microbial inoculants. They boost agricultural yields while conserving the lithosphere and benefiting the biosphere by preventing soil, water, and air pollution and eutrophication. The use of PGPR containing strains of beneficial microorganisms is one strategy used in sustainable farming to reduce issues brought on by overuse of mineral fertilizers (Mishra and Dash, 2014).
TYPES OF PGPR
PGPR can improve nutrient absorption and promote overall plant growth. These PGPR form symbiotic relationships with plants or fix atmospheric nitrogen, making it accessible to plants in a usable form. There are diverse types of PGPR, each serving a specific purpose on crops:
Nitrogen-fixing PGPR: These contain nitrogen-fixing bacteria, such as Rhizobium, Azotobacter, and Azospirillum, which convert atmospheric nitrogen into a form that plants can utilize.
Phosphorus-solubilizing PGPR: These consist of bacteria and fungi, such as Bacillus and Pseudomonas, which help solubilize insoluble phosphates in the soil, making phosphorus more available to plants.
Potassium-mobilizing PGPR: These contain microorganisms that enhance the availability of potassium in the soil, improving the plant's ability to absorb this essential nutrient.
Symbiotic PGPR: These involve the use of mycorrhizal fungi or nitrogen-fixing bacteria that form symbiotic relationships with plant roots, aiding in nutrient absorption and overall plant health.
For ease of understanding, we categorized them based on their mode of action in the following sub-headings which are as follows:
Nutrient enhancement: PGPR that can fix atmospheric nitrogen and solubilize phosphorus, making these essential nutrients available to plants which could enhance soil fertility, healthier plant growth and their crop yields.
Reduced dependency on fertilizers: The application of PGPR reduces the reliance on synthetic or chemical fertilizers, which can have detrimental effects on soil health and the environment over time. Biofertilizers can offer a more sustainable and eco-friendlier alternative by promoting natural nutrient cycles.
Environmental sustainability: PGPR contributes to the conservation of soil, water, and air quality. These helps maintain the balance of ecosystems by minimizing the negative impacts associated with the excessive use of chemical fertilizers, such as soil degradation, water pollution, and greenhouse gas emissions.
Promotion of microbial activity: PGPR introduces beneficial microorganisms into the soil, fostering a healthy microbial community around plant root ecosystem. These beneficial microbes play a crucial role in nutrient cycling, disease control attributes, and overall healthy soil ecosystem functioning.
Biodiversity conservation: By promoting sustainable farming practices, PGPR contributes to the conservation of biodiversity. Reduced dependence on chemical inputs helps maintain a more balanced and diverse ecosystem in and around agricultural areas.
Improved soil health: The activities of PGPR can enhance soil structure by promoting the formation of aggregates. This improves water retention, aeration, and drainage in the soil, leading to better overall soil quality.
Cost-effectiveness: In modern agriculture, PGPR can be cost-effective for farmers for sustainable agriculture. Though the initial investment might be higher, the reduced need for chemical fertilizers, coupled with their improved soil fertility and crop yields, can lead to overall economic returns to farmers.
Plant tolerance: Some PGPR that can enhance a plant's resistance to various environmental stresses, such as drought, salinity, and certain diseases. This can contribute to increased crop resilience and adaptability to changing climatic conditions.
PGPR MARKET TRENDS
Some of the biofertilizers market trends are:
Growing demand for organic agriculture: The increasing global demand for organic produce has been a significant driver for the PGPR market. Consumers are more conscious of the environmental impact of conventional farming practices, leading to a shift towards organic and sustainable agriculture.
Government initiatives and regulations: Most of the governments around the globe have been promoting the use of PGPR through subsidies, incentives, and regulations. This support is aimed at encouraging farmers to adopt eco-friendly and sustainable agricultural practices.
Advancements in PGPR strains: Current research and development in microbiology have led to the identification and isolation of more effective microbial strains for PGPR production. Improved strains contribute to better nutrient availability and overall plant health.
Education in PGPR: Farmers and agricultural practitioners are becoming more aware of the benefits of PGPR. Educational programs and initiatives by agricultural extension services contribute to the dissemination of knowledge regarding the advantages of using biobased PGPR.
Compatibility with conventional chemical fertilizers: PGPR are increasingly being integrated into conventional fertilizer practices. This integrated approach helps optimize nutrient management, reduce dependency on chemical fertilizers, and enhance soil health.
Bio-technological innovations in PGPR production: Bio-technological innovations in the production of PGPR have led to increased efficiency and cost-effectiveness. Improved manufacturing processes and packaging methods contribute to the wider adoption of PGPR.
Focus on Nitrogen-fixing PGPR: Nitrogen-fixing bacteria, such as Rhizobium and Azotobacter, are gaining prominence due to their role in enhancing nitrogen availability in the soil. This is particularly crucial, as nitrogen is a vital nutrient for plant growth.
Market opportunities and partnerships: The PGPR industry has witnessed various mergers, acquisitions, and strategic partnerships. Such collaborations help companies broaden their product portfolios, enhance research capabilities, and expand their market reach.
PGPR in climate resilience: The environmental benefits of PGPR, such as reduced greenhouse gas emissions and improved soil structure, align with global sustainability goals. As climate change concerns grow, the emphasis on sustainable agricultural practices is likely to drive the adoption of PGPR.
APPLICATION OF PGPR FOR SUSTAINABLE AGRICULTURE
Seed application: PGPR can be applied directly to seeds before planting. This mode ensures that the beneficial microorganisms establish contact with the emerging roots in the preliminary stages of plant growth.
Soil Application: PGPR can be mixed with soil before sowing seeds or transplanting seedlings. This allows the PGPR to colonize the root zone, promoting a symbiotic relationship with the plants.
Seedling drench or dip: Seedlings can be immersed in a PGPR solution before transplanting. This method ensures that the young plants are inoculated with beneficial microorganisms, fostering a favorable environment for root development. A liquid PGPR solution can be poured or sprayed directly onto the roots of established plants. This method facilitates the direct application of beneficial PGPR to the root system, enhancing nutrient uptake.
Foliar spray: PGPR can be applied as a foliar spray onto the leaves of plants. This spay allows for the absorption of nutrients through the stomata on the leaf surface, providing a supplemental nutrient source to support plant growth.
Drip irrigation: PGPR can be injected into irrigation systems for controlled and uniform distribution. This system ensures that plants receive a consistent supply of beneficial microorganisms along with water.
Integration with low dose fertigation: This method involves the integration of PGPR into conventional low dose fertilizer application practices. It combines fertilization with irrigation, allowing for the simultaneous delivery of nutrients and beneficial microorganisms to the root zone.
Application with compost: PGPR can be incorporated into composting processes. Adding beneficial microorganisms during composting enhances the microbial diversity and nutrient content of the final compost, which can then be applied to the soil.
Application in hydroponics: In soilless growing systems like aquaponics and hydroponics, PGPR can be introduced to the nutrient solution. This application provides a source of beneficial microorganisms to support plant growth in these controlled environments.
The formulation of PGPR can consist of one or more distinct microbial strains, such as AM, Bacillus, Pseudomonas, Rhizobium, Acetobacter, Azotobacter, Rhizobium and other PGPR strains. The PGPR can be classified into different groups.
NITROGEN FIXING PGPR
An element necessary for life, nitrogen (N) is a part of amino acids, nucleic acids, chlorophyll, and the energy-transfer molecule adenosine triphosphate (ATP). Organic N is one of the main sources of nitrogen in soils. It must undergo microbial mineralization i.e., through processes like nitrification and ammonification, to be transformed into inorganic N that is available to plants. The main reservoir of nitrogen, however, is found in the atmosphere as N2, which is only made available via biological nitrogen-fixation and is not utilized by plants directly. The energy-intensive process of biological nitrogen fixation involves the conversion of atmospheric N2 to ammonia (NH3) by the enzyme nitrogenase. This is a readily assimilable form for both plants and microorganisms. Nitrogenases are present in a relatively few diverse classes of microorganisms known as diazotrophs (N2-fixing), which includes free-living bacteria, symbiotic bacteria, and archaea. Multiple bacterial strains, such as aerobic (Azotobacter), anaerobic (Clostridium), facultatively anaerobic (Klebsiella), or heterotrophs; anoxygenic (Rhodobacter) or oxygenic (Anabaena) phototrophs and chemolithotrophs (Leptospirillum ferrooxidans), can carry out nitrogen fixation with varying physiological requirements. Diazotrophs inhabit a variety of soil and aquatic habitats. They can establish symbiotic relationships with cyanobacteria, and actinorhizal associations with woody plants, symbiotic relationships with legumes that result in the production of root nodules. BNF permits the replenishment of the total nitrogen contentand fixed nitrogen controls crop productivity and growth.
The Rhizobiaceae family of bacteria, which includes Rhizobium, Bradyrhizobium, Sinorhizobium, Azorhizobium, Mesorhizobium, and Sinorhizobium, are the most studied group of symbiotic N2-fixing organisms. By forming nodules on the roots or stems of legumes, they can form symbiotic associations. The benefit of these nodules for N2-fixation is that bacteroids' nitrogenases are protected from atmosphere O2. Given that oxygen is a negative regulator of nif gene expression and inhibits nitrogenase activity, the quantity of oxygen plays a significant role in controlling the amount of N that is fixed. A considerable increase in plant biomass and grain yield was seen in a variety of legume crops after efficient rhizobia strains were inoculated. Rhizobium strain inoculation of soybeans resulted in an overall increase in growth and yield components, including the number of branches yielding pods per plant, the total number of pods per plant, and the number of seeds per pod (Mfilinge et al., 2014). Bourion et al., (2018) reported that when R. leguminosarum strains inoculated into pea and lentil seeds, nodulation, shoot/root weight, and pea seed production were all increased. Legume plants are less reliant on chemical fertilizers than non-leguminous plants because of the nitrogen fixation process that Rhizobium performs.
Several heterotrophic free-living diazotrophic microorganisms, including cyanobacteria, Azotobacter sp. and Azospirillum sp., can fix atmospheric N2 in the rhizosphere and bulk soil. In non-legume crops, free-living diazotrophs play a crucial role in N acquisition. enhanced yields of many crops, including tomato, eggplant, pepper, cotton, sunflower, carrot, oak, and sugar beet, have been observed, for instance, along with enhanced yields of rice, corn, and wheat (Garcha and Maan, 2017). According to Jain et al., (2021) findings, seven of the 24 Azotobacter strains that were identified demonstrated phosphate solubilization, while allthe 24 strains produced ammonia, siderophores, HCN, and IAA. Certain strains of cyanobacteria have been shown to boost the growth and yield of radish, maize, tomato, cotton, barley, and oats. Several legumes showed significantly enhanced nodule number, nitrogen fixation, plant biomass, and total nitrogen contents when compared to uninoculated plants or a single Rhizobium inoculation. The most well-known free-living diazotrophs, Azospirillum species have been proved to improve N availability and acquisition in addition to performing a number other PGP activity. Azospirillum species associates with plant roots and synthesize various compounds involved in plant growth promotion, for instance IAA, gibberellins and cytokinin (Pereg et al., 2016) are shown in Table 1.
Table 1. Types of nitrogen fixing PGPR strains.
Types of biofertilizer
PGPR strain used
Plant growth promoting activity
Effect on plant productivity parameters
Nitrogen-fixing
bacteria
Azotobacter sp. strain Avi2 (MCC 3432)
Nitrogen fixation, production of
IAA, siderophore
Improved vegetative and reproductive growth in
rice
Azotobacter chroococcum, A. vinelandii
Nitrogen fixation, P
solubilization, production of
NH3, HCN, IAA
Increased shoot and root length, leaf and rootnumber, chlorophyll content of maize
Herbaspirillum seropedicae (strain
ZAE94)
Nitrogen fixation, production of
siderophore, IAA
Enhanced mineral uptake, increased diameter,
weight, length of ear, number of grains/rows, cob
weight and grain yield
Bradyrhizobium sp
Nitrogen fixation, siderophore
and IAA production, P
solubilisation
Increased vegetative parameters and seed yield in
mung bean
Rhizobium meliloti
Nitrogen fixation, production of
siderophore and chitinase
Increased growth and yield, pods quality and better
use of nitrogen in peanut
PHOSPHATE SOLUBILIZING PGPR
One of the most crucial nutrients for plants is phosphorus, which influences every biological function either directly or indirectly. P is essential for all major metabolic activities in plants, including respiration, energy transmission, photosynthesis, signal transduction, and molecular biosynthesis. Although soils contain a significant amount of P in both organic and inorganic forms, the availability of P is one of the primary factors limiting plant growth. This is because plants can absorb P from the soil solution as orthophosphate ions H2PO− 4 and HPO42−, but the majority of soil P is in insoluble state and is therefore unavailable (Soumare et al., 2020). Phosphorous biofertilizers include both phosphate solubilizers and mobilizers. Phosphate-solubilizing microorganisms (PSM) produce and release several chemicals that help solubilize inorganic P, such as tricalcium phosphate, hydroxyapatite, and rock phosphate. The release of organic acids, hydroxyl ions and CO2 are one method. This lowers the pH of the soil, directly dissolving the insoluble phosphates and causing PO42− to be exchanged for acid ions (Wei et al., 2018).Moreover, microbes could produce chelating substances, which can be used to release related soluble phosphates by capturing and mobilizing cations from various insoluble phosphates, such as Ca+2, Al+3, and Fe+3. PSM reduce the need for mineral P fertilizer inputs, which when used excessively can have detrimental effects on the environment, such as eutrophication of freshwater bodies, by raising P bioavailability. The most investigated P solubilizers belong to genera Pseudomonas, Bacillus, Rhizobium, Enterobacter, Penicillium, and Aspergillus (Anand et al., 2016; Riaz et al., 2020).
It has been reported that phosphorus in soil is solubilizedby a variety of microorganisms, such as Bacillus, Pseudomonas, Achromobacter, Brevibacterium, Erwinia sp., Flavobacterium sp., Micrococcus sp., Corynebacterium, Xanthomonas sp., Nostoc, Rhodococcus sp., Serratia phosphaticum, Acytonema, Calothrix brauna, and Tolypothrixceylonica, Bacillus, Pseudomonas, Achromobacter, Brevibacterium, Burkholderia, Sarcina sp., and Scytonema. Like this, it has been observed that a variety of fungi, such as Alternaria sp., Aspergillus sp., Cephalosporium sp., Fusarium sp., Rhodotrulaminuta, Saccharomyces cerevisiae, Torula thermophila, Paeciliomyces, Penicillium, and Sclerotium rolfisi, exhibit phosphate solubilization (Sindhu et al., 2014; Santoyo et al., 2021).
When chickpea was inoculated with PSB, the number of nodules per plant, shoot length, number of pods per plant, and grain production increased by 23%, 13%, 17%, and 15%, respectively. In addition, the stability of the soil aggregate increased by 37%, and 2.35 times more phosphorous was liberated from the rock phosphate. Additionally, the protein, phosphorus and nitrogen contents of chickpea and straw were increased to 16%, 42%, and 11%, respectively (Ditta et al., 2018).
In addition, mycorrhiza is essential for the cycling of nutrients, mobilization of phosphorus and the growth of microbial biomass. Native arbuscular mycorrhizae (AM) are primarily found in soil, where they colonize plant roots and promote plant growth. The availability of phosphorous increases abruptly when mycorrhiza is introduced into low-phosphorus soil. The primary function of arbuscular mycorrhizal fungi, which are found in nature in symbiotic relationships with cereals and horticultural plants, is to increase the bioavailability of phosphorus. Plants with mycorrhizal associations out compete those without in terms of utilizing soil nutrients because fungi have long enough hyphae to reach distant soils that plant roots are unable to access. Gigaspora, Acaulospora, Glomus, Scutellospora, and other genera are commonly used as biofertilizers (Pandey et al., 2019). Mycorrhiza has a beneficial effect on the structure of the soil as well. Improved plant health, soil productivity, soil aggregate stability and increased agricultural production are among the benefits of mycorrhizal inoculation.
POTASSIUM SOLUBILIZING PGPR
Potassium (K) is a key inorganic cation found in the cytoplasm of plants, and it is a crucial macronutrient for plants. It is involved in many basic metabolic processes, including protein synthesis, photosynthesis, and cell structure and function. In addition to being one of the most prevalent elements on Earth, potassium is the second most abundant nutrient in soil after nitrogen. Nonetheless, approximately 98% of soil K is found in a non-exchangeable state within the crystal structures of feldspar and mica (such as biotite and muscovite). Only 0.1–0.2% of the total is in the soil solution and immediately available for plant absorption, with the remaining 1-2% adsorbed onto organic matter and clay particles (Mitter et al., 2021). By solubilizing K, microorganisms can make it more accessible to plants, which is an important part of the K cycle. The release of organic acids, such as tartaric, citric, oxalic, gluconic, lactic, and malic acid, is the most well-known process of microbial K solubilization. (Macik et al., 2020).
Due to their diversity and complexity in terms of phylogeny, microbes interact with plants through a variety of mechanisms, such as symbiosis, parasitism, amensalism and neutralism (Fig. 1). These microorganisms are collectively referred to as the plant microbiome because they are dependent on photosynthesis in plants and reciprocate by influencing plant growth. Since mycorrhizal fungi and PGPR have been proposed to be essential for both nutrient availability in the soil and alleviation of stress, these microorganisms have grown to be key components in the efficient operation of the soil ecosystem.
Certain phytohormones, or plant growth regulators, are synthesized by bacteria and plants in very small amounts. These hormones affect a variety of physiological processes, such as cell division, development, gene expression, and stress responses, as well as the shape, flowering, senescence, and seed growth. PGPR either produce hormones or change the hormone concentration in the plant in response to biotic and abiotic stressors. Many microbial species have been identified as producing gibberellins and cytokinin’s, which are bioactive hormones that influence a variety of developmental processes, including fruit setting, blooming, stem elongation, root hair growth, and seed germination (Khanet al., 2020). It was discovered that bacteria that produce siderophores that are essential for both biocontrol and growth stimulation. Siderophores are produced by Pseudomonas, Aeromonas, Azotobacter, Bacillus, Rhizobium, Azadirachta, Streptomyces, Burkholderia and Serratia.
Arbuscular mycorrhizal fungi (AMF) are obligate symbionts which can create symbiotic associations with around 80% of land plant species, including crops used in agriculture. AMF provides various advantages, including improving the uptake of water and mineral nutrients (such as P, N, S, Cu, and Zn) by their host plants in return for a carbon source. The extraradical AMF mycelium expands the amount of soil that is explored by entering smaller soil holes in addition to extending well beyond the rhizosphere. This is especially critical for the uptake of less mobile nutrients like P, and AMF is widely recognized for its capacity to improve P uptake, particularly in soils lacking in P (Fig. 2.).

ADVANTAGES AND LIMITATIONS OF PGPR
PGPR have the potential to significantly improve environmental sustainability by lowering soil, water, and air pollution brought on by chemical fertilizers. It improves soil fertility, boosts microbial biodiversity, aids in plant survival under a variety of stressful situations, and controls plant disease. Nevertheless, a few restrictions on PGPR prevented their widespread use and commercialization. The main obstacles to industrial-scale production and commercialization include the use of fertilizers with lower nutrient contents than chemical fertilizers, variations in or deviations from their efficacy when stored and exposed to external conditions such as light, temperature, and humidity, inappropriate carrier materials, inappropriate microbial strains, a lack of quality assurance, inexperienced and inefficient staff, low volume production, improper inoculation techniques, mutated strains, and market availabilityare the major limitations in the industrial scale production and commercialization of biofertilizers (Fig. 3).
CASE STUDIES
The experiment conducted during summer of 2018 at ICAR-Indian Agricultural Research Institute, New Delhi to study the effect of crop establishment practices and microbial inoculation on growth and nutrient uptake by summer green gram (Vigna radiata (L.) Wilczek). Results indicated that zero tillage with 2.5 t ha-1 residue produced significantly higher growth parameters like crop growth rate (CGR) and net assimilation rate (NAR) at 20, 35 and at 50 DAS, grain (968.6 kg ha-1) and (2415.3 kg ha-1) stover yield. The same treatment showed significantly higher uptake of N, P and K in grain and stover over other two treatments. Seed inoculation with Rhizobium+PSB+AM fungi was significantly superior to other two treatments about growth parameters like CGR and NAR at 20, 35 and 50 DAS, grain yield, stover yield and uptake of N, P and K in grain and stover of summer green gram. Highly positive correlations were recorded between grain yield and yield attributes like grain yield and pods/plant, grain yield and grains/ pod, grain yield and test weight of summer green gram (Naragund, et al., 2020).
Shome et al., 2022 conducted following a completely randomized design to explore the impact of nitrogen fixing bacteria and PSB on the growth, yield, and quality attributes of soybean alongside soil nutrient availability using Rhizobium japonicum and Pseudomonas striata. The experiment consisted of two factors, R. japonicum (100% N, R. japonicum alone or with 50% N and control) and P. striata (100% P, P. striata with 75% P and control). Results revealed a significant influence of interaction on seed N, yield, protein, oil, and nodules of soybean. Microbial inoculants with or without N and P fertilizers produced a statistically similar yield as 100% N and P. Furthermore, R. japonicum and P. striata along with 50% N and 75% P increased 7% protein and 19% oil than 100% N and P. R. japonicum enhanced soil N content and P. striata improved soil phosphorus availability. Overall, R. japonicum and P. striata inoculation with 50% N and 75% P can potentially improve the yield and the quality of soybean and soil nutrient conditions.
Shilpa et al., (2022) in her studies based on the utilization of carbon sources, thirty isolates were identified as Azospirillum brasiliense, and twenty isolates were identified as A. amazonense. Selected isolates were tested for their antifungal activity against Exserohilum turcicum causal agent of turcicum blight of maize under in vitro condition and antifungal activity of isolates was determined by the decreased diameter of fungal growth compared to the diameter of the control fungal growth. The isolate ACDAZS-35 showed the highest percentage inhibition of 85.56%. The isolate ACD-15 at different nitrogen levels showed a significant increase in number of leaves over the uninoculated control. It was found that maximum number of leaves, SPAD values, N content was observed in ACDAZS-9 inoculated with 50% N application at 30 and 60 DAS. The present study revealed that the selected Azospirillum inoculants could increase the growth of maize plants and can save 25% of inorganic nitrogen application.
Azolla has been exploited widely as biofertilizer for rice and some crop plants, and it fixes atmospheric nitrogen due to the presence of a symbiotic Cyanobacterium and Anabaena azollae. It has several other uses such as food, feed, biogas producer and hyper accumulator of heavy metals, etc. The use of biofertilizers like Azolla not only increased the crop productivity but also improved the long-term soil fertility. These multi-dimensional uses of Azolla-Anabaena system would be ideal and environment friendly in sustainable agriculture. The current study critically reviewed the Azolla’s potential capacity to be used in sustainable agriculture (Santhiya and Jeeva, 2022).
A study conducted by Reddy and Rahe (1989) monitored the populations of a marked strain of the B-2 isolate of B. subtilis (B-2) in the rhizospheres of onion seedlings grown from bacterized seeds in muck soil at various pH, moisture and temperature regimes for 14 weeks. Irrespective of regime, populations of B-2 in onion rhizospheres declined rapidly within the first 14 days after seeding, from 4.8×106 cfu per seed on day zero to a mean of 1.1×103 cfu per plant at day 14, and less rapidly during the next 12 weeks following seeding.
Various formulations are available in the market viz., carrier based, oil and liquid based formulations. This carrier must be nontoxic for plants and microbes, physically and chemically stable, cost-effective, biodegradable and able to maintain humidity, and ensure cell viability (Balla et al., 2022 and Mącik et al., 2020). Peat-based inoculants have historically dominated the commercial biofertilizer market owing to the substantial surface area of peat, its excellent water retention properties, and the conducive environment it provides for metabolic activity and cell proliferation during storage. Furthermore, due to their carbon dioxide trapping capacity, if peat is used as a carrier, its mitigation effect on climate change will be reversed when the captured CO2 is released again. In addition to their negative environmental impact on peat-rich ecosystems (John et al., 2011, Balla et al., 2022, Gewin, 2020). Thus, there has been a shift towards the development of new carriers from both organic materials (such as compost, biogas slurry, crushed corn cobs, biochar, peat, etc.) and inorganic substances (including zeolite, perlite, lignite, or vermiculite) (Thomas et al., 2019, John et al., 2011).
Field trials conducted over two growing seasons indicated that none of the 19 PGPR strains, applied as seed treatments at two concentrations, significantly reduced late leaf spot disease. The same chemical elicitors tested in the greenhouse, including BABA, did not elicit significant disease protection. Some combinations of four PGPR and BABA significantly reduced the disease at one but not at two sample times. Collectively, these results suggest that late leaf spot resistance in peanut is not systemically inducible in the same manner as is resistance to diseases in other crops by PGPR and chemical inducers Zhang et al., (2001).
A carotenoid producing halotolerant PGPR Dietzia natronolimnaea STR1 protecting wheat plants from salt stress by modulating the transcriptional machinery responsible for salinity tolerance in plants. The expression studies confirmed the involvement of ABA-signalling cascade, as TaABARE and TaOPR1 were upregulated in PGPR inoculated plants leading to induction of TaMYB and TaWRKY expression followed by stimulation of expression of a plethora of stress related genes. Enhanced expression of TaST, a salt stress-induced gene, associated with promoting salinity tolerance was observed in PGPR inoculated plants in comparison to uninoculated control plants. (Bharti et al., 2016).
ADVANCEMENTS IN PGPR
In the recent years, several advancements have developed to enhance the nutrients and growth dynamics of soil and plant for the betterment of crop production such as specialized microorganism are used for the inoculation of plants, best technologies are chosen, good carrier material for inoculum purposes. Furthermore, to reduce the cost of biofertilizer and for the sustainable production, current approaches are the artificial selection of microbiomes, mixed inoculants, PGPR as biofertilizers, use of biofilms as biofertilizers and Nano-biofertilizers.
MOLECULAR MECHANISM OF N2 FIXATION BY PGPR
Rhizobacteria is a nitrogen fixing bacteria that has developed symbiotic association and form nodules with the leguminous plants. They contain specialized genes known as Nif that carry out N2 fixation and are present in free existing as well as symbiotic bacteria. They also contain nitrogenase that includes reductase with Fe and Mo as cofactor and dinitrogenase reductase enzyme with iron cofactor. Dinitrogenase causes the reduction of nitrogen to ammonia by electrons that are facilitated by dinitrogenase reductase (Olanrewaju et al., 2017). Among Nif gene there are several regulatory transporters who contributed to the activation and formation of enzyme also regulate the synthesis of iron proteins. For the expression of Nif gene oxygen is an inhibitor but needed for the respiration of Rhizobium species. Oxygen causes the activation of the enzyme by binding with leg hemoglobin of bacteria with great affinity. Hence, to continue the process of nitrogen fixation accurately, significant amount of oxygen is facilitated to the rhizobacterium for respiration Basu et al., 2021). Genetic engineering is the easiest method to complete this process by transforming bacterial hemoglobin bounded with oxygen into bacteria. In result, respiratory rate of bacterial cells is increased up to several folds which in turn increase the rate nitrogen fixation to 68% (Gouda et al., 2018). Bacteria such as Azorhizobium and Rhizobium are symbiotic bacteria that fixes nitrogen by forming stem modules. They also enhance the growth of plant by synthesizing large amount of indole acetic acid. An efficient nitrogen fixer Bradyrhizobium when inoculated with the seeds of Mucuna, then it will enhance the potassium, Nitrogen, carbon content and phosphorus in the soil. Azotobacters are free living bacteria that produce different types of Phytohormones and vitamin B complex to promote pathogen resistance as well as growth of the crops (Sayyed et al., 2019). Azolla and Cyanobacteria are efficiently using as a biofertilizer because these species are useful as a source of nitrogen in the production of rice. Mainly Azolla is employed in the field before the cultivation of rice as a green fertilizer and supply essential amount of nutrients (Olanrewaju et al., 2017).
DEVELOPMENT OF MIXED PGPR
The use of mixture of bacterial strains instead of one is considered an important approach as different mechanism of action provide more potential for the use of biofertilizer. Mixed inoculant contains microbial consortia instead of single microbial strain that facilitate combined biofertilization effect and help plants to promote better uptake of nutrients. Recent studies have investigated a combined effect of co-inoculant containing Arbuscular mycorrhizal fungi (AMF) and rhizobacteria for the enhanced growth of the leguminous plant (Igiehon and Babalola, 2017). Co-inoculant of microbial specie allow wide range of biofertilizer efficiency and reliability for the fixation of nitrogen, phosphate solubilization and siderophore production and balanced growth and nutrition as compared to a single inoculant (Viswanath, 2019).
ENCAPSULATION AND LYOPHILISATION TECHNIQUE
Encapsulation technology is one of the current emerging techniques which allow the controlled release of biofertilizer into the environment. This technique is widely used to develop microbial species having regulating composition and structure characteristics (Schoebitz et al., 2013). Among this technology there is an advanced subfield of microencapsulation that works on the mechanism of constructing a protective covering such as capsule or a shell across an active molecule that could increase crop productivity and shelf life of biofertilizer. This type of formulation is more advantageous in term of slow and sustained release into soil and provide protection against environmental stress to biofertilizer (Çakmakçı, 2019).
Lyophilisation is the freeze-drying method for the preservation and storage of microorganism. It is widely used technique that helps to make a laboratory formulation of microorganisms. Lyophilisation is an efficient way to increase the survival rate of bacteria and storage for longer period; thus, the lyophilized microbial organisms can be used directly in combination with a suitable carrier in the field. This lyophilized structure can be used applied directly or a suitable carrier material can be used with it (Hernández et al., 2006).
NANO-PGPR
Nano-biofertilizer is the combined use of nano technology and biofertilizer for the more efficient and enhanced productivity of crops. The synergistic effect of nanomaterial and microbial fertilizer facilitates the maintenance of soil moisture and uptake of vital nutrients for plant (Kumari and Singh, 2019). Major issues of other fertilizers, such as instability on field due to fluctuation in environmental condition, temperature and change of pH, poor shelf life, reduced microbial strains, only short-term efficiency and need of large amount of fertilizer to cover wide area etc can lead to poor growth and provide less yield (Çakmakçı, 2019). The use of nano biofertilizer provide essential benefit to farmers in term of good field performance, low cost, less expenses, and provide more yield (Mala et al., 2017). Nano-biofertilizer are not only ecofriendly with the environment but also produce best quality crops by enhancing the efficiency of indigenous microbial population through utilization of essential nutrients such as potassium, nitrogen, and phosphorus. This will also stimulate the activity of microbial enzymes also contribute to the fertility of soil.
NANO-ENCAPSULATION TECHNOLOGY
Nano-encapsulation is the coating of biofertilizer at a nanoscale with the help of a nanomaterial. PGPR containing biofertilizers can be protected by using nanoencapsulation technology (Golbashy et al., 2017). This is a versatile technology which facilitates increased shelf life, provide controlled release of PGPR also allow controlled diffusion by biofertilizer. Nanomaterial is used for the nanoencapsulation of the nutrients and organic compounds such as zeolite, polymers and chitosan that facilitate the gradual and controlled bioavailability of nutrients to plants (Qureshi et al., 2018). Nano biofertilizer act synergistically with the microbial species or AMF inoculants through the enrichment of soil via bioavailability of nutrients that is carried out by several mechanisms such as fixation of nitrogen by enhanced activity of PGPR, phytohormone production, solubilization of phosphorus, and production of chelating agents that can chelate the elements and release them for plant uptake (Mala et al., 2017).
BIOFILM PGPR
Biofilms are the microbial communities that are be either biotic or abiotic and embed with some biological component that facilitates sustainability and structural characteristic to the biofilm (Junaid and Khan, 2018). Microbial species work together by making linkage with each other through the process of quorum sensing. The main objective for the development of biofilm biofertilizer is to overcome the problems of biotic and a biotic component of soil and to make the indigenous microbial population more resistive against pathogens and inhibitors (Parween et al., 2017). Biofilm is either synthesized on the plant surface or inside the plant and becomes advantageous for the fixation of nitrogen by PGPR and enhance the productivity of crops. Biofilm also help in Zn solubilization and production of chelating agents (Igiehon and Babalola, 2017). Biofilms provide a way to transfer the genetic material and organic nutrients between microbial species in a community. As compared to a biofilm comprising of single microbial species, multiple microbial strains in a biofilm are investigated to be more resistant and sustainable approach. For instance, the uptake of nutrient and tolerance to environmental stress is significantly increased for bacterial-fungal biofilm as compared to those microorganisms that are single or do not form biofilm (Velmourougane et al., 2017). Biofilm biofertilizer provide resistance to crops against heavy metal, drought, stress, pH, temperature, and attack of pathogen. Biofilm biofertilizer also help in the preservation of land and lessen harmful consequences on the human health. In addition to this, it also increases the productivity of the crops under different environmental conditions and facilitates longer shelf life to farmers (Mitter et al., 2021).
PGPR MARKET SIZE AND TRENDS
The global PGPR market was valued at 2.8 billion in 2022 and is projected to reach USD 5.2 billion by 2028, growing at a CAGR of 10.9% during the study period. The market size is estimated to be valued at USD 3.1 billion in 2023 (Fig. 4). Through natural processes such as nitrogen fixation; phosphate solubilization; and the production of compounds that stimulate growth, PGPR provides nutrients to plants. They boost soil organic matter and aid in restoring the soil's natural nutrition cycle. By using PGPR, growers can increase the sustainability and health of the soil and cultivate healthy crops. Only bacteria and other biological elements that are not damaging to the environment are present in PGPR. As a result, they contribute to reducing pollution brought on by agricultural activities, particularly soil pollution.
SMART PGPR IN SUSTAINABLE AGRICULTURE
The first Green Revolution (1965–1985) and the post Green Revolution period (1985≈2005) resulted in tremendous yield growth in staple crops, helping to feed the world despite large population increases and decreasing land availability for agriculture. The classical breeding methods and advancements in genomics, genome sequencing, transgenics, genome editing, and related breeding methodologies such as marker-assisted selection and incorporation of phenomics, data analytics, and artificial intelligence for the rapid development of climate-resilient crops. The utilisation of newer methods to understand essential genetic and molecular components of the multiple plant microbe interactions (PM) is required for their application. The use of clustered regularly interspaced short palindromic repeats (CRISPR)/Cas-mediated genome editing (GE) techniques to investigate PM interactions is of tremendous interest. The implementation of GE techniques to boost the ability of microorganisms or plants for agronomic trait development will be enabled by a comprehensive understanding of PM interactions. As a result, utilising the plant microbiota is a viable strategy for the next Secondary Green Revolution for meeting food demand.
MODE OF ACTION OF PGPR
The mode of action of PGPRs on crop plants can be direct or indirect. Direct mechanisms include biofertilization (nitrogen (N2) fixation, production of plant hormones and siderophores (SDs), phosphorus (P) solubilization, etc.), root growth stimulation, rhizoremediation, and the control of plant stress. Additionally, PGPRs produce metabolites, such as indole-3-acetic acid (IAA), 1-aminocyclopropane-1-carboxylate-deaminase (ACC-deaminase), phosphate solubilizing enzyme (PSE), and SDs. They can also indirectly enhance plant growth by reducing the detrimental effects of phytopathogens through induced systemic resistance (ISR) and the production of antimicrobial compounds, such as bacteriocin, zwittermicin, fengycin, chitinase, and cell wall-degrading enzymes (Fig. 5)
FORMULATIONS OF PGPR
From many years in the field of agriculture application, biofertilizers has made higher achievements for the sustainable production of crops and establish more resilient conditions for the growth of microbes. But the use of conventional fertilizer formulation led to several pathogenic problems and reduced viability of microbial species during the field application (Çakmakçı, 2019). Therefore, the advanced suitable formulation for making a bioinoculant is essential so that the beneficial indigenous soil population can effectively develop under plant roots. For that purpose, microorganism need such type of protective formulation strategies that are effective at the target site and provide benefit to farmers (Stamenković et al., 2018). Formulation needs to be stable for longer period and should remain viable during the process of synthesis, packaging, transport and on field application (Mitter et al., 2021).
There are different PGPR formulations related to its recent approaches such as metabolite formulations, liquid formulations, solid carrier-based formulations, synthetic polymer-based formulations, granular formulations, powered formulations. The bioinoculant formulation is the uniform mixture of selected beneficial strain with a suitable carrier that can provide stabilization and protection of strain during transport and storage. The carrier is the vehicle of living latent microbes that provides protection and supportive niche to the microbial community. A good bioformulation should be effective, non-polluting, readily biodegradable with high water retention capacity and sufficient shelf life. There are still lots of potential of using innovative technologies towards improving the capability of bioinoculant, but it needs better integration of ideas and utilizing them for sustainable agriculture.
REGULATORY CONSTRAINS OF PGPR
Although the PGPR technology is a low cost, eco-friendly technology, several constraints limit the application or implementation of the technology. The constraints may be environmental, technological, infrastructural, financial, human resources, unawareness, quality, marketing, etc. The different constraints, in one way or another, affect the technique at production or marketing or usage. The guideline limits itself to only biofertilizers or biofertilizers containing carrier based (solid or liquid) living microorganisms which colonize the rhizosphere or the interior of the plant and promotes growth by increasing the supply or availability of primary nutrient and/or growth stimulus to the target crop, when applied to seed, plant surfaces, or soil. Throughout the guidelines of many countries, biofertilizers will refer to microbial biofertilizers. Common data requirements and equivalent registration systems pave the way for harmonisation of biofertilizer registration either within a region or across the entire continent. Flexibility is provided in this framework for registration upon the extent of regional integration. Our understanding is that we do not need any registration for PGPR based biofertilizers. Our Asian PGPR Society for Sustainable Agriculture (www.asianpgpr.com) is being developing guidelines for registration of PGPR as biofertilizers around the globe.
SPECIFIC RESEARCH NEEDS ON PGPR
1. Selection of effective and competitive multi-functional PGPR for a variety of crops.
2. Quality control system to produce PGPR and their application in the field, to ensure and explore the benefits of plant-microorganism symbiosis.
3. Study of microbial persistence of PGPR in soil environments under stressful conditions
4. Agronomic, soil, and economic evaluation of PGPR for diverse agricultural production systems.
5. Transferring technological know-how on PGPR production to the industrial level and for optimum formulation.
6. Establishment of "Bio-fertilizer Act" and strict regulation for quality control in markets and application.
CONCLUSIONS
PGPR offer a sustainable alternative, harnessing microbial power to enhance soil fertility and plant growth without adverse effects. Understanding soil ecology and microbial dynamics is crucial for optimizing PGPR efficacy. Leveraging genetic and biotechnological advancements can lead to improved formulations and production methods, paving the way for start-ups to explore this promising field. The application of inorganic chemical fertilizers was thus significantly reduced to 30-50%. This helps in the realization of environmentally friendly and sustainable agriculture.
Excess nutrients are accumulated in soils, particularly P because of over application of chemical fertilizers by farmers during intensive agricultural practices. Hence, major research focus should be on the production of efficient and sustainable bio-fertilizers for crop plants, wherein inorganic fertilizer application can be reduced significantly to avoid further pollution problems. In view of overcoming this bottleneck, it will be necessary to undertake short-term, medium, and long-term research, in which soil microbiologists, agronomists, plant breeders, plant pathologists, and even nutritionists and economists must work together.
Further research on the nano-encapsulation, multiple biofilms formation and mixed microbial PGPR are continuing to engineer disease resistance indigenous microorganism and plants. Future goal of PGPR is to develop innovative and eco-friendly agricultural techniques that are cheaper than use of chemical fertilizers. Thanks to the recent advances in molecular systems PGPR and the development of new-generation technology platforms for high efficacy microbial ecology studies, a collaborative, cross-disciplinary research effort, from ecological to bioprocess and metabolic engineering, will pave the way for predictable microbial diversity manipulation and new microbial-driven solutions to tackle the current environmental challenges we are facing at the nexus of food, water and energy.
REFERENCES
Anand, K. U., B. A. Kumari and M.A. Mallick. 2016. Phosphate solubilizing microbes: an effective and alternative approach as biofertilizers. Int. J.Pharm. Pharm. Sci. 8(2): 37-40.
Asoegwu, C. R, C. G. Awuchi, K. C. Nelson, C. G. Orji, O. U. Nwosu and U. C. Egbufor. 2020. A review on the role of biofertilizers in reducing soil pollution and increasing soil nutrients. Himalayan Journal of Agriculture. 1(1): 34-8.
Adeleke, R. A., A. R. Raimi, A. Roopnarain and S. M. Mokubedi. 2019. Status and prospects of bacterial inoculants for sustainable management of agroecosystems. p. 137-72. In B. Giri, R. Prasad, Q. S. Wu and A. Varma (eds.), Biofertilizers Sustain. Agric. Environ. Soil Biol. Springer, Cham, 10.1007/978-3-030-18933-47.
Bala, A., N. Karanja, M. Murwira, L. Lwimbi, R. Abaidoo and K. Giller. 2011. Production and use of Rhizobial inoculants in Africa, Www.N2 Africa.Org. 1–22.
Balla, A., A. Silini, H. Cherif-Silini, A. Chenari Bouket, F. N. Alenezi and L. Belbahri. 2022. Recent Advances in Encapsulation Techniques of Plant Growth-Promoting Microorganisms and Their Prospects in the Sustainable Agriculture. Appl. Sci. 12 (18), 9020. https://doi.org/10.3390/app12189020.
Basu, A., P. Prasad, S. N. Das, S. Kalam, R. Z. Sayyed, M. S. Reddy and E.l.H. Enshasy. 2021. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: recent developments, constraints, and prospects. Sustainability. 13(3): 1140. https://doi.org/10.3390/su13031140.
Bharti, N., S. S. Pandey, B. Deepti, V. K. Patel and A. Kalra. 2016. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci Rep. 6: 34768. https://doi.org/10.1038/srep34768.
Bourion, V., K. Heulin-Gotty, V. Aubert, P. Tisseyre, M. Chabert-Martinello, M. Pervent, C. Delaitre, D. Vile, M. Siol, G. Duc and B. Brunel 2018. Co-inoculation of a pea core-collection with diverse rhizobial strains shows competitiveness for nodulation and efficiency of nitrogen fixation are distinct traits in the interaction. Frontiers in Plant Science. 8: 290940. doi: 10.3389/fpls.2017.02249.
Çakmakçı, R. 2019. A Review of Biological Fertilizers Current use, New Approaches, and Future Perspectives. International Journal of Innovative Studies in Sciences and Engineering Technology. 5(7): 83-92
Chansa-ngavej, C. and S. Assavavipapan. 2007. Forecasting of rhizobial biofertilizer technology using maturity mapping. Proceedings of the Int. Colloq. Bus. Manag., Bangkok, Academia. P. 1-32
Ditta, A., M. Imtiaz, S. Mehmood, M. S. Rizwan, F. Mubeen, O. Aziz, Z. Qian, R. Ijaz and S. Tu. 2018. Rock phosphate-enriched organic fertilizer with phosphate-solubilizing microorganisms improves nodulation, growth, and yield of legumes. Communications in Soil Science and Plant Analysis. 49(21): 2715-25.
Garcha, S. and P. K. Maan. 2017. Biological nitrogen fixation in cereals crops: a bacterial perspective. Advances in Soil Microbiology: Recent Trends and Future Prospects: Soil Microbe Plant Interaction. 6(2): 127-51.
Graham, P. H., and C. P. Vance. 2003. Update on legume utilization legumes: importance and constraints to greater use. Plant Physiol. 131. p. 872-877, 10.1104/pp.017004.872.
García-Fraile, P., E. Menéndez and R. Rivas. 2015. Role of bacterial biofertilizers in agriculture and forestry. AIMS Bioeng. 2. p. 183-205, 10.3934/bioeng.2015.3.183.
Gewin, V. 2020. How peat could protect the planet. Nature. 578: 204–208.
Gouda, S., R. G. Kerry, G. Das, S. Paramithiotis, H. S. Shin and J. K. Patra. 2018. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiological Research, 206: 131–140. https://doi.org/10.1016/j.micres.2017.08.016.
Hernández, A, F. Weekers, J. Mena, C. Borroto and P. Thonart. 2006. Freeze-drying of the biocontrol agent Tsukamurella paurometabola C-924 predicted stability of formulated powders. Industrial Biotechnology. 2(3): 209-212.
Hegde, G. M., S. Aditya, D. Wangdi and B. K. Chetri. 2022. Mycoremediation: A Natural Solution for Unnatural Problems. In: V. R. Rajpal, I. Singh and S. S. Navi, S.S. (eds) Fungal diversity, ecology and control management. Fungal Biology. Springer, Singapore. https://doi.org/10.1007/978-981-16-8877-517.
Igiehon, N. O. and O. O. Babalola. 2017. Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Applied Microbiology and Biotechnology. 101(12): 4871–4881. https://doi.org/10.1007/s00253-017-8344-z.
Jain, D., J. Sharma, G. Kaur, A. A. Bhojiya, S. Chauhan, V. Sharma, A. Suman, S. R. Mohanty and E. Maharjan. 2021. Phenetic and molecular diversity of nitrogen fixating plant growth promoting Azotobacter isolated from semiarid regions of India. Bio.Med. Research International. 5(3): 221-29.
John, R. P., R. D. Tyagi, S. K. Brar, R. Y. Surampalli and D. Prévost. 2011. Bio-encapsulation of microbial cells for targeted agricultural delivery. Crit. Rev. Biotechnol. 31: 211-226.
Junaid, P. M. and F. Khan. 2018). Biofilm: A future generation biofertilizer. Indian Journal of Applied and Pure Biology. 33(2): 131-134.
Khan, N., A. Bano, S. Ali and M. A. Babar. 2020. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regulation. 90: 189-203.
Kumari, R., and D. P. Singh. 2019. Nano-biofertilizer: An emerging eco-friendly approach for sustainable agriculture. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences. 1-9.
Mala, R., R. Celsia Arul Selvaraj, V. Barathi Sundaram, R. Blessina Siva Shanmuga Rajan and U. Maheswari Gurusamy. 2017. Evaluation of nano structured slow-release fertilizer on the soil fertility, yield and nutritional profile of vigna radiata. Recent Patents on Nanotechnology. 11(1): 50-62.
Lawal, T. E. and O. O. Babalola. 2014. Relevance of biofertilizers to agriculture J. Hum. Ecol. 47: 35-43, 10.1080/09709274.2014.11906737.
Macik, M., A. Gryta and M. Frac. 2020. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Advances in Agronomy. 16 (2): 31-87.
Mfilinge, A., K. Mtei and P. Ndakidemi. 2014. Effect of Rhizobium inoculation and supplementation with phosphorus and potassium on growth and total leaf chlorophyll (Chl) content of bush bean Phaseolus vulgaris, L. Agricultural Sciences. 5: 1413-1426
Effect of Rhizobium Inoculation and Supplementa-
tion with Phosphorus and Potassium on Growth and Total Leaf Chlorophyll (Chl) Content of Bush Bean Phaseolus vulgaris, L.
Mishra, P. and D. Dash. 2014. Rejuvenation of biofertilizer for sustainable agriculture and economic development. Consilience. 1(11): 41-61.
Mitter, E. K., M. Tosi, D. Obregón, K. E. Dunfield and J. J. Germida. 2021. Rethinking crop nutrition in times of modern microbiology: innovative biofertilizer technologies. Frontiers in Sustainable Food Systems. 5: https://doi.org/10.3389/fsufs.2021.606815.
Malusà, E., L. J. Sas-Paszt and Ciesielska. 2012. Technologies for beneficial microorganisms inocula used as biofertilizers Sci. World J. 1-12: 10.1100/2012/491206.
Naragund, R., Y. V. Singh, R. S. Bana, A. K. Choudhary, J. Pranita, K. Praveen, and S. Andhimansu. 2020. Influence of crop establishment practices and microbial inoculants on plant growth and nutrient uptake of summer green gram (Vigna radiata) Annals of Plant and Soil Research. 22(1): 55-59.
Olanrewaju, O. S., B. R. Glick and O. O. Babalola. 2017. Mechanisms of action of plant growth promoting bacteria. World Journal of Microbiology and Biotechnology. 33(11): 197. https://doi.org/10.1007/s11274-017-2364-9.
Pandey, D., H. K. Kehri, I. Zoomi O. Akhtar and A. K. Singh. 2019. Mycorrhizal fungi: Biodiversity, ecological significance, and industrial applications. Recent Advancement in White Biotechnology Through Fungi: Diversity and Enzymes Perspectives. 1: 181-99.
Pereg L, L. E. de-Bashan and Y. Bashan. 2016. Assessment of affinity and specificity of Azospirillum for plants. Plant and Soil. 39(9): 389-414.
Riaz, U., S. M. Mehdi, S. Iqbal, H. I. Khalid, A. A. Qadir, W. Anum, M. Ahmad and G. Murtaza. 2020. Bio-fertilizers: eco-friendly approach for plant and soil environment. Romberger, J. A. and P. Mikola. 1966. International Review of Forestry Research. Academic Press, New York. 10.2307/forhis/10.2.20-a.
Reddy, M. S. and H. Singh. 2011. Plant growth-promoting rhizobacteria (pgpr) for sustainable agriculture. Field evaluation of P solubilising bacteria on the growth and yield of wheat and maize grown in rock phosphate amended soils - Proceedings of the 2nd Asian PGPR Conference August 21-24. Beijing, P. R. China.
Reddy, M. S. and J. E. Rahe. 1989. Growth effects associated with seed bacterization not correlated with populations of Bacillus subtilis inoculant in onion seedling rhizospheres. Soil Biology and Biochemistry. 21(3): 373-378.
Strijdom, B.W. 1998. South African studies on biological nitrogen-fixing systems and the exploitation of the nodule bacterium-legume symbiosis. S. Afr. J. Sci. 94: 11-23.
Santoyo, G., P. Guzmán-Guzmán, F. I. Parra-Cota, S. D. Santos-Villalobos, M. D. Orozco-Mosqueda and B. R. Glick. 2021, Plant growth stimulation by microbial consortia. Agronomy. 11(2): 219. https://doi.org/10.3390/agronomy11020219.
Sindhu, S. S., M. Phour, S. R. Choudhary and D. Chaudhary. 2014. Phosphorus cycling: prospects of using rhizosphere microorganisms for improving phosphorus nutrition of plants. In: Parmar, N., Singh, A. (eds). P. 199-237. Geomicrobiology and Biogeochemistry. Soil Biology, vol 39. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-41837-2_11 199-237.
Soumare, A., K. Boubekri, K. Lyamlouli, M. Hafidi, Y. Ouhdouch and L. Kouisni. 2020. From isolation of phosphate solubilizing microbes to their formulation and use as biofertilizers: status and needs. Frontiers in bioengineering and biotechnology. 7: 425-35.
Sanhthosh, K. R. G. and M. Sandeep. 2020. Biofertilizers role in major pulse crops of India International Journal of All Research Education and Scientific Methods (IJARESM), ISSN: 2455-6211 Volume 8, Issue 11.
Shome, S., A. Barman and Zakaria M. Solaiman. 2022. Rhizobium and Phosphate Solubilizing Bacteria Influence the Soil Nutrient Availability, Growth, Yield, and Quality of Soybean (Bangaldesh) Agriculture. 12: 1136. https://doi.org/10.3390/ agriculture 12081136.
Santhiya, B. and S. Jeeva. 2022. Azolla as a source of biofertilizer for sustainable crop production. Journal of Xi’an Shiyou University, Natural Science Edition ISSN: v1673-064X. 18 (11) 598-605.
Schoebitz, M., M. D. López and A. Roldán. 2013. Bioencapsulation of microbial inoculants for better soil–plant fertilization. A review. Agronomy for Sustainable Development. 33(4): 751-765.
Sayyed, R. Z., M. S. Reddy and S. Antonius. (Eds.). 2019. Plant growth promoting rhizobacteria (PGPR): Prospects for Sustainable Agriculture. Springer Singapore.
Shilpa, H., V. Navi, G. P. Santhosh and G. M. Hegde. 2022. Isolation, identification of Azospirillum and its inoculation effects on maize (Zea mays L.) J. Farm Sci. 35(3): 370-380.
Stamenković, S., V. Beškoski, I. Karabegović, M. Lazić and N. Nikolić. 2018. Microbial fertilizers: A comprehensive review of current findings and future perspectives. Spanish Journal of Agricultural Research. 16(1), e09R01. https://doi.org/10.5424/sjar/2018161-12117.
Thomas, L. and I. Singh. 2019. Microbial biofertilizers: types and applications. pp. 109-135. In Giri, B., R. Prasad, Q. Wu and A. Varma (Eds.), Biofertilizers Sustain. Agric. Environ., Vol 55, Springer Nature, Switzerland AG. 10.1007/978-3-030-18933-41.
Viswanath, B. 2019. Recent Developments in Applied Microbiology and Biochemistry. p.363-373.
Velmourougane, K., R. Prasanna and A. K. Saxena. 2017. Agriculturally important microbial biofilms: present status and prospects. Journal of Basic Microbiology: 57(7): 548-573.
Wei, Y., Y. Zhao, M. Shi, Z. Cao, Q. Lu, T. Yang, Y. Fan and Z. Wei. 2018. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresource Technology. 24 (7):190-99.
Zhang, S., M. S. Reddy, N. Kokalis-Burelle, L. W. Wells, S. P. Nightengale and J. W. Kloepper. 2001. Lack of induced systemic resistance in peanut to late leaf spot disease by plant growth-promoting rhizobacteria and chemical elicitors. Plant Dis. 85: 879-884.